
C 2 -
CYSTEINYL LEUKOTRIENE RECEPTOR 2 ACTIVATION MEDIATES POST-MYOCARDIAL ISCHEMIA/REPERFUSION INJURY INFLAMMATORY PROCESSES BY NATHAN CHEN NI A THESIS SUBMITTED TO THE GRADUATE PROGRAM IN PHYSIOLOGY IN CONFORMITY WITH THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY QUEEN'S UNIVERSITY KINGSTON, ONTARIO, CANADA SEPTEMBER, 2013 COPYRIGHT © NATHAN CHEN NI, 2013 Abstract Myocardial infarction (MI) is primarily caused by blockade of the coronary circulation, resulting in ischemic insult. The only available remedy is reperfusion, which induces oxidative stress and activates inflammatory responses at the site of injury. Cysteinyl leukotrienes (cysLTs) are potent pro-inflammatory mediators that exert their effects through two classical receptors: cysLT receptor 1 (CysLT 1 R) and cysLT receptor 2 (CysLT 2 R), the latter of which is prevalent in the heart and circulatory system and has been implicated in cardiovascular disease. However, although endothelial CysLT 2 R overexpression exacerbates MI damage and induces vascular hyperpermeability, understanding of CysLT 2 R activation-induced mechanisms is poor, as isolating CysLT 2 R-specific effects has proven difficult due to a lack of appropriate pharmacological agents. We investigate herein the role of CysLT 2 R activation in myocardial ischemia/reperfusion injury. We have characterized a novel CysLT 2 R-selective antagonist BayCysLT 2 in both in vitro and in vivo systems, and establish that CysLT 2 R-selective antagonism attenuates exacerbated MI injury, adhesion molecule gene regulation, and myocardial neutrophil presence observed in CysLT 2 R overexpressing (EC) mice. We also examined effects of CysLT 2 R antagonism in long-term cardiac remodeling post-myocardial infarction, and found that blockade of CysLT 2 R post-reperfusion, regardless of whether CysLT 2 R is overexpressed or not, elicits a mild pathological cardiac hypertrophic response despite mitigating infarction damage to the apical ventricular wall. Finally, we created a novel mouse model (EC/KO) that expresses CysLT 2 R predominantly in vascular endothelium in order to identify tissue-specific mechanisms of CysLT 2 R activation. Surprisingly, MI injury was attenuated in EC/KO mice, indicating that both endothelial and non-endothelial CysLT 2 R ii expression subsets have roles in mediating infarction injury. Indeed, EC/KO mice demonstrated hyperpermeability in cremaster venules only when leukotrienes are applied, in contrast to EC mice. In addition, endothelial CysLT 2 R activation facilitates leukocyte transmigration, whereas non-endothelial CysLT 2 Rs regulate basal rolling leukocyte flux in microvasculature. Although much work remains to be done, the characterization of a CysLT 2 R-selective antagonist provides a vital tool for CysLT 2 R research moving forward, and our investigation of CysLT 2 R activation reveal the existence of a complicated and multi-faceted pathway resulting in activation of proinflammatory mechanisms. iii Co-Authorship Nathan C. Ni, as part of a collaboration with Dr. Colin D. Funk, conceived all hypotheses examined in the current work. Unless otherwise stated, Nathan C. Ni designed and performed experiments, analyzed results, and disseminated the collective works presented for publication. Dong Yan, as part of a collaboration with Dr. Derek A. Pratt, synthesized BayCysLT 2 for Chapter 2 and generated the data presented in Figures 2-1, 2-2, 2-3, and 2-5, as well as the figures themselves. Dr. Alma Barajas-Espinosa assisted with the myeloperoxidase assay for Chapter 2. Laurel L. Ballantyne performed the LAD ligation surgery for Chapters 2, 3, and 4, the cremaster muscle surgery for Chapter 4, assisted with the vascular ear permeability assay for Chapter 2, and conducted the ultrasound measurements under the advisement of Nathan C. Ni for Chapter 3. Tim St. Amand provided the transfected HEK-293 cells and assisted with the calcium imaging experiments for Chapter 2. Jeffrey D. Mewburn performed the cremaster muscle surgery and provided technical assistance for intravital microscopy image capture and analysis for Chapter 4. Chapter 2 has been published in the Journal of Pharmacology and Experimental Therapeutics (Volume 339 (3), pages 768-78, Dec 2011). iv Acknowledgements First and foremost, my gratitude goes to my supervisor, Dr. Colin D. Funk, for his support and mentorship throughout the course of this project, for teaching me much about not only the academic side of science, but also about lab management and how to cultivate a successful research environment. This degree would not be possible without him. My gratitude goes out as well to my committee members Dr. Shetuan Zhang, Dr. Christopher A. Ward, Dr. Donald H. Maurice, and Dr. Janos Filep, for their invaluable advice and commentary. To the Funk lab, past and present, you have my gratitude for providing a unique learning and working environment throughout these four years. Thank you for being there through the ups and downs, the good and the bad. I would like to give special thanks to Mr. Tim “LoveGame” St. Amand, Ms. Laurel “Poker Face” Ballantyne, Ms. Crystal “Just Dance” McCracken, Dr. Alma “Alejandro” Barajas Espinosa, and Mr. Dong “The Edge of Glory” Yan for their camaraderie, advice, support, and friendship over the years. See, I did learn something from that blasted radio after all. I would like to offer my sincere thanks to my parents for all of their effort and love over the years, for their sweat – and occasionally even blood. Although I can never repay all that they have given me, I can do my best to at least provide the next generation the same opportunities. Finally, to my wife Samantha, who has been with me through thick and thin, sickness and health, and to the ends of the world: ES ERGO SUM. v Table of Contents ABSTRACT................................................................................................................................... ii CO-AUTHORSHIP ..................................................................................................................... iv ACKNOWLEDGEMENTS ......................................................................................................... v LIST OF FIGURES AND ILLUSTRATIONS........................................................................... x LIST OF TABLES ....................................................................................................................... xi LIST OF ABBREVIATIONS .................................................................................................... xii CHAPTER 1: GENERAL INTRODUCTION ..................................................................................... 1 1.1. MYOCARDIAL INFARCTION ................................................................................................... 2 1.1.1. Cause and Prevalence .................................................................................................. 2 1.1.2. Ischemic Injury ............................................................................................................. 3 1.1.3. Reperfusion Injury ........................................................................................................ 4 1.1.4. Post-Reperfusion Inflammation Cascades.................................................................... 6 1.2. LEUKOTRIENES...................................................................................................................... 9 1.2.1. Leukotriene Synthesis Pathway .................................................................................. 10 1.2.2. Leukotriene Transcellular Biosynthesis...................................................................... 16 1.2.3. Leukotriene B 4 ............................................................................................................ 18 1.2.4. Cysteinyl Leukotrienes................................................................................................ 20 1.3. CYSTEINYL LEUKOTRIENE RECEPTORS ............................................................................... 22 1.3.1. Cysteinyl Leukotriene Receptor 1 ............................................................................... 22 1.3.2. Cysteinyl Leukotriene Receptor 2 ............................................................................... 25 1.3.3. Other Cysteinyl Leukotriene Receptors ...................................................................... 27 1.4. LEUKOTRIENE INVOLVEMENT IN PATHOLOGY .................................................................... 30 1.4.1. Asthma/Respiratory Diseases ..................................................................................... 30 1.4.2. Cancer......................................................................................................................... 32 1.4.3. Ischemic/Cardiovascular Disease .............................................................................. 34 1.5. RESEARCH OBJECTIVES AND HYPOTHESES ......................................................................... 39 CHAPTER 2: A SELECTIVE CYSTEINYL LEUKOTRIENE RECEPTOR 2 ANTAGONIST BLOCKS MYOCARDIAL ISCHEMIA/REPERFUSION INJURY AND VASCULAR PERMEABILITY IN MICE ......... 41 2.1. ABSTRACT ........................................................................................................................... 42 2.2. INTRODUCTION.................................................................................................................... 44 2.3. MATERIALS AND METHODS ................................................................................................ 47 vi 2.3.1. BayCysLT 2 Synthesis .................................................................................................. 47 2.3.2. β-Galactosidase-β-Arrestin Assay .............................................................................. 51 2.3.3. Measurement of Agonist-Induced Intracellular Calcium Mobilization...................... 51 2.3.4. Animal Models ............................................................................................................ 53 2.3.5. Vascular Ear Permeability Assay ............................................................................... 53 2.3.6. Ischemia/Reperfusion Surgical Model........................................................................ 54 2.3.7. Analysis of Infarct Volume.......................................................................................... 55 2.3.8. RNA Extraction and Quantitative Real-Time PCR..................................................... 56 2.3.9. Myeloperoxidase Assay............................................................................................... 57 2.3.10. Statistical Analysis.................................................................................................... 59 2.4. RESULTS ............................................................................................................................. 60 2.4.1. Synthesis and chemistry .............................................................................................. 60 2.4.2. BayCysLT 2 is more potent than Bay-u9773................................................................ 60 2.4.3. BayCysLT 2 is selective for CysLT 2 R in vitro and in vivo ........................................... 62 2.4.4. BayCysLT 2 attenuates exacerbation of myocardial damage in EC mice ................... 66 2.4.5. Upregulation of adhesion molecule gene expression is attenuated by BayCysLT 2 .... 68 2.5. DISCUSSION......................................................................................................................... 72 2.6. ACKNOWLEDGEMENTS ........................................................................................................ 76 CHAPTER 3:. LONG-TERM MYOCARDIAL REMODELING POST-ISCHEMIA/REPERFUSION INJURY IS MILDLY IMPAIRED BY CYSLT 2 R ANTAGONISM. ...................................................................... 77 3.1. ABSTRACT ........................................................................................................................... 78 3.2. INTRODUCTION.................................................................................................................... 79 3.3. MATERIALS AND METHODS ................................................................................................ 82 3.3.1. Mouse Models ............................................................................................................. 82 3.3.2. Myocardial Infarction Induction ................................................................................ 82 3.3.3. Echocardiography ...................................................................................................... 83 3.3.4. Statistical Analysis...................................................................................................... 85 3.4. RESULTS ............................................................................................................................. 86 3.4.1. Age-related weight gain post-ischemia/reperfusion is unaltered by overexpression of endothelial CysLT 2 R or by BayCysLT 2 treatment. ............................................................... 86 3.4.2. Post-ischemia/reperfusion pathological cardiac hypertrophy is exacerbated in animals treated with BayCysLT 2 . ......................................................................................... 86 3.4.3. Mice treated with BayCysLT 2 show mildly decreased cardiac function. ................... 93 vii 3.4.4. Left ventricular volume and mass are increased in BayCysLT 2 -treated mice postischemia/reperfusion injury. ................................................................................................. 93 3.5. DISCUSSION......................................................................................................................... 97 3.5.1. CysLT 2 R blockade post-I/R injury promotes pathological cardiac hypertrophy. ...... 97 3.5.2. Why does CysLT 2 R overexpression not result in a worse long-term outcome? ......... 98 3.5.3. Why does CysLT 2 R blockade result in an apparently worse long-term outcome?..... 99 3.6. ACKNOWLEDGEMENTS ...................................................................................................... 101 CHAPTER 4: MULTIPLE-SITE ACTIVATION OF THE CYSTEINYL LEUKOTRIENE RECEPTOR 2 IS REQUIRED FOR EXACERBATION OF ISCHEMIA/REPERFUSION INJURY ...................................... 102 4.1. ABSTRACT ......................................................................................................................... 103 4.2. INTRODUCTION.................................................................................................................. 105 4.3. MATERIALS AND METHODS .............................................................................................. 108 4.3.1. Mouse Models ........................................................................................................... 108 4.3.2. Myocardial Ischemia/Reperfusion............................................................................ 108 4.3.3. Analysis of Infarct Volume........................................................................................ 110 4.3.4. Intravital Microscopy of the Cremaster Vasculature ............................................... 111 4.3.5. Analysis of Vascular Permeability............................................................................ 112 4.3.6. Analysis of Vascular Leukocytes............................................................................... 112 4.3.7. Complete Blood Count.............................................................................................. 113 4.3.8. Statistical Analysis.................................................................................................... 113 4.4. RESULTS ........................................................................................................................... 115 4.4.1. Endothelium-targeted CysLT 2 R overexpression does not exacerbate infarction damage in the absence of endogenous CysLT 2 R ................................................................ 115 4.4.2. Vascular permeability responses in EC/KO mice differ from those in EC mice ...... 115 4.4.3. Genetic knockout of endogenous CysLT 2 R reduces basal rolling leukocyte flux in cremaster vasculature. ........................................................................................................ 121 4.4.4. Endothelial-targeted CysLT 2 R overexpression results in diminishing leukocyte flux in cremaster vasculature following leukotriene stimulation ................................................... 121 4.4.5. Cysteinyl leukotriene stimulation results in increased leukocyte slow rolling fraction in cremaster vasculature in mice expressing endothelial CysLT 2 R. .................................. 123 4.5. DISCUSSION....................................................................................................................... 128 4.5.1. Endothelial CysLT 2 R is not the sole mediator of CysLT 2 R-mediated ischemia/reperfusion injury exacerbation .......................................................................... 128 4.5.2. Various tissue-specific CysLT 2 R niches play divergent, yet synergistic, roles in mediating exacerbation of CysLT 2 R-mediated ischemia/reperfusion injury...................... 132 viii CHAPTER 5: GENERAL DISCUSSION....................................................................................... 136 5.1. CHARACTERIZATION OF A NOVEL CYSLT 2 R-SELECTIVE ANTAGONIST ............................ 137 5.2. IDENTIFICATION OF CYSLT 2 R-MEDIATED MECHANISMS POST-MYOCARDIAL INFARCTION ................................................................................................................................................. 139 5.2.1. CysLT 2 R is the main cysteinyl leukotriene receptor involved in myocardial infarction responses ............................................................................................................................. 139 5.2.2. CysLT 2 R activation results in promotion of cell adhesion and vascular hyperpermeability ............................................................................................................... 141 5.2.3. CysLT 2 R activation on different cell types mediate distinct and separate responses ............................................................................................................................................ 143 5.2.4. The CysLT 2 R picture is far from complete ............................................................... 148 5.3. CYSLT 2 R ANTAGONISM POST-REPERFUSION IMPEDES LONG-TERM VENTRICULAR REMODELING ........................................................................................................................... 152 5.4. POTENTIAL FOR THE CYSLT 2 R AS A THERAPEUTIC TARGET............................................. 154 REFERENCES.......................................................................................................................... 156 APPENDIX................................................................................................................................ 193 FORMULAE USED TO CALCULATE CARDIAC FUNCTION FROM ECHOCARDIOGRAPHY IMAGES . 193 ix List of Figures and Illustrations Figure 1-1: Leukotriene synthesis pathway. ................................................................................. 12 Figure 2-1: Chemical synthesis of BayCysLT 2 . ........................................................................... 61 Figure 2-2: BayCysLT2 is a more potent antagonist of the human CysLT2 receptor than Bay-u9773 .. 63 Figure 2-3: BayCysLT 2 selectively antagonizes the human CysLT2 receptor calcium responses.... 64 Figure 2-4: BayCysLT 2 is ≥500-fold more selective for CysLT 2 R than CysLT 1 R. .................... 65 Figure 2-5: BayCysLT 2 is a potent selective antagonist at the CysLT 2 receptor in vivo. ............ 67 Figure 2-6: BayCysLT 2 attenuates CysLT 2 R-mediated exacerbation of myocardial ischemia/ reperfusion injury in mice. .................................................................................................... 69 Figure 2-7: BayCysLT 2 attenuates indirect measures of leukocyte recruitment in ischemia/ reperfusion injury-induced diapedesis. ................................................................................. 71 Figure 3-1: Echocardiography imaging and left ventricular dimensional measurement methodology... 84 Figure 3-2: Representative images of left ventricular remodeling post-myocardial infarction.... 89 Figure 4-1: Myocardial injury is not elevated in EC/KO mice following ischemia/reperfusion..... 117 Figure 4-2: Vascular permeability post-surgical intervention (“basal”) is elevated in EC mice, but not in EC/KO mice.............................................................................................................. 119 Figure 4-3: CysLT 2 R activation mediates vascular hyperpermeability. .................................... 120 Figure 4-4: Basal rolling leukocyte flux is mediated by endogenous CysLT 2 R. ....................... 122 Figure 4-5: Rolling leukocyte flux diminishes in mice expressing elevated endothelial CysLT 2 R post-leukotriene stimulation................................................................................................ 124 Figure 4-6: Rolling leukocyte velocity distributions in mice before and after cysteinyl leukotriene stimulation........................................................................................................................... 126 Figure 4-7: Rolling leukocyte velocity medians in mice before and after cysteinyl leukotriene stimulation........................................................................................................................... 127 Figure 4-8: Proposed schematic for CysLT 2 R inflammatory response modulation................... 134 x List of Tables Table 2-1: Primers used in qualitative real-time PCRs................................................................. 58 Table 3-1: Body weights of mice prior to and following ischemia/reperfusion injury at time of echocardiography.................................................................................................................. 87 Table 3-2: Left ventricular wall measurements of mice post-I/R injury. ..................................... 90 Table 3-3: Left ventricular dimensions of mice post-I/R injury. .................................................. 91 Table 3-4: Apex wall thickness measurements of mice post-I/R injury....................................... 92 Table 3-5: Evaluation of functional measurements in mice post-I/R injury................................. 94 Table 3-6: Left ventricular volume and mass measurements in mice post-I/R injury.................. 96 Table 4-1: CysLT 2 R expression profile in featured mouse genotypes. ...................................... 109 xi List of Abbreviations 5-HpETE aa ADP AMP ANOVA ATP BAL BayCysLT 2 Bay-u9773 BLT 1 R BLT 2 R bpm cAMP CD CO COX cPLA 2 α CVD CysLT CysLT 1 R CysLT 2 R CysLT E R DiP DMSO EC EC/KO EC 50 EF ERK EtOAc FITC FLAP FS GAPDH GGT GM-CSF GPCR 5-hydroperoxyeicosatetraenoic acid amino acid adenosine diphosphate adenosine monophosphate analysis of variance adenosine triphosphate bronchoalveolar lavage 3-[[(3-carboxycyclohexyl)amino]carbonyl]-4-[3-[4-(4-phenoxybutoxy) phenyl]propoxy]-benzoic acid 4-(((1R,2E,4E,6Z,9Z)-1-((1S)-4-carboxy-1-hydroxybutyl)-2,4,6,9pentadecatetraen-1-yl)thio)benzoic acid Leukotriene B 4 receptor 1 Leukotriene B 4 receptor 2 beats per minute cyclic adenosine monophosphate cluster of differentiation cardiac output cycloxygenase cytosolic phospholipase A 2 α cardiovascular disease cysteinyl leukotriene cysteinyl leukotriene receptor 1 cysteinyl leukotriene receptor 2 cysteinyl leukotriene receptor E dipeptidase dimethyl sulfoxide endothelial cell-targeted CysLT 2 R-overexpressing transgenic mouse model endothelial cell-"restricted" CysLT 2 R-overexpressing transgenic mouse model half maximal effective concentration ejection fraction extracellular signal-regulated kinase ethyl acetate fluorescein isothiocyanate 5-lipoxygenase-activating protein fractional shortening glyceraldehyde 3-phosphate dehydrogenase γ-glutamyl transferase granulocyte/macrophage colony-stimulating factor G protein-coupled receptor xii GPR17 GPR99 GSK HAMI3379 HEK HUVEC I/R IC 50 ICAM IFN IL IVS KO LAD LFA LO LPS LT LTA 4 H LTC 4 S LV LVID LVOD LVPW MAPK MAPKAPK MCP M-CSF mGST MI MIP MK571 MMP MPO MRP NAD(P)H NMLTC 4 NMR P2Y 12 G protein-coupled receptor 17 G protein-coupled receptor 99 glycogen synthase kinase 3-[[(3-carboxycyclohexyl)amino]carbonyl]-4-[3-[4-[4-(cyclohexyloxy) butoxy]phenyl]propoxy]-benzoic acid human embryonic kidney human umbilical vein endothelial cell ischemia/reperfusion half maximal inhibitory concentration intercellular adhesion molecule interferon interleukin interventricular septum knockout left anterior descending coronary artery lymphocyte function-associated antigen lipoxygenase lipopolysaccharide leukotriene leukotriene A 4 hydrolase leukotriene C 4 synthase left ventricle left ventricular internal dimension left ventricular outer dimension left ventricular posterior wall mitogen-activated protein kinases MAP kinase-activated protein kinase monocyte chemotactic protein macrophage colony-stimulating factor microsomal glutathione-S-transferase myocardial infarction macrophage inflammatory protein (E)-3-[[[3-[2-(7-chloro-2-quinolinyl)ethenyl]phenyl][[3-(dimethylamino)-3oxopropyl]thio]methyl]thio]-propanoic acid matrix metalloproteinases myeloperoxidase multidrug resistance-associated protein Nicotinamide adenine dinucleotide (phosphate) N-methyl leukotriene C 4 nuclear magnetic resonance purinergic receptor P2Y, G-protein coupled, 12 xiii PAF PBS PG PI3K PKA PKC RLU ROS SEM SRS SRS-A SV TEM TGF THF TLC TLR TNF UDP VCAM platelet activating factor phosphate-buffered saline prostaglandin phosphatidylinositide 3-kinase protein kinase A protein kinase C relative light unit reactive oxygen species standard error of the mean slow reacting substance slow reacting substance of anaphylaxis stroke volume Tie2-expressing monocyte transforming growth factor tetrahydrofuran thin layer chromatography toll-like receptor tumour necrosis factor uridine diphosphate vascular cell adhesion molecule xiv Chapter 1: General Introduction Cardiovascular disease (CVD) is a major problem in modern society: it resulted in over 800,000 deaths (25% more than cancer) in the United States alone in 2008. In addition, CVD related cases cost the United States health system an estimated ~300 billion dollars per year. Globally, cardiovascular disease strikes primarily industrialized nations, but is prevalent in Asia, Europe, and the Americas, making it a global health issue. Further exacerbating the problem is that cardiovascular disease risk factors, primarily obesity (present in 68% of the United States adult population), hypertension (33.5%), and diabetes mellitus (11.1%, plus an additional 37% with prediabetes) are on the rise, and medical costs relating to cardiovascular heart disease are projected to increase by ~200% in the next twenty years. While the death rate has plateaued in recent years, the number of CVD related operations performed in the United States increased 22% from 1999 to 2009, indicating an overall increase in disease prevalence (Heidenreich et al., 2011; Roger et al., 2012). Treatment options for cardiovascular disease are largely post-hoc, focusing on limiting damage, where possible, rather than directly attacking underlying mechanisms responsible for pathogenesis, which is problematic as the heart possesses relatively limited regenerative properties. Preventative therapy mainly centers around lifestyle changes in order to remove CVD risk factors or use of pharmaceutical agents to suppress them. Herein, we examine the involvement of leukotrienes, pro-inflammatory lipid mediators, in myocardial ischemia/reperfusion injury, and focus in particular on activation of the cysteinyl leukotriene receptor 2 (cysLT 2 R) as a mediator of leukotriene-triggered pathways during and following ischemia/reperfusion injury. We seek to identify potential therapeutic targets for the treatment of 1 cardiovascular disease by elucidating the molecular, genetic, and cellular mechanisms underlying CysLT 2 R-mediated ischemia/reperfusion injury size modulation. 1.1. Myocardial Infarction 1.1.1. Cause and Prevalence Myocardial infarction (MI), commonly referred to as a “heart attack” (although it is not the sole condition covered by that blanket vernacular), is a situation where myocardial blood supply is interrupted, leading to oxygen and nutrient deprivation of the myocardium. If left untreated, MI leads to myocardial necrosis and loss of cardiac function. The main cause of coronary arterial blockage is physical disruption of a lipid and macrophage filled atherosclerotic plaque, resulting in the formation of a thrombus which partially or fully occludes the vascular lumen (Davies et al., 1976; Van de Werf et al., 2008). Long-term treatment usually revolves around removal or circumvention of the obstruction (i.e. stenting or coronary bypass surgery). Acute treatment is surgical (i.e. angioplasty) or pharmacological (i.e. thrombolysis) intervention to restore blood flow (Van de Werf et al., 2008). Timely reperfusion of the myocardium is not without its caveats, as a phenomenon termed “reperfusion injury” has been noted for many years - and will be explored later in this introduction (Yellon and Hausenloy, 2007). Of all deaths due to cardiovascular disease in the United States, 50% were attributed to coronary heart disease (including myocardial infarction), and the projected number of coronary heart disease sufferers is projected to rise by as many as 8 million people in 20 years. Myocardial 2 infarction sufferers are also more susceptible to cardiovascular heart disease (whether stroke, heart failure, or another infarction) than the general population, and show a 1.5 to 15-fold increased chance of illness or death following a first myocardial infarction (Roger et al., 2012). Myocardial infarction pathogenesis is extremely complicated, but known risk factors include age, gender, hypercholesterolemia/hyperlipidemia, obesity, diabetes mellitus, hypertension, smoking, and stress (Graham et al., 2007; Nabel and Braunwald, 2012; Roger et al., 2012; Smith et al., 2006). 1.1.2. Ischemic Injury Myocardial ischemia, as noted previously, is characterized by a coronary arterial blockage resulting in oxygen and nutrient deficiency in the affected myocardium. Upon onset of ischemia, mitochondrial oxidative phosphorylation ceases within seconds, resulting in the loss of the primary ATP production pathway in the cell and a compensatory increase in anaerobic glycolysis. Increased glycolysis results in H+ and lactate accumulation, creating a state of intracellular acidosis (Buja, 2005; Kloner and Jennings, 2001). These metabolic alterations result in ATP and creatine phosphate depletion, as well as adenine nucleotide pool degradation. The latter occurs because ADP is formed via the actions of various cellular ATPases much more rapidly than the phosphorylation of ADP to ATP by anaerobic glycolysis (Rovetto et al., 1975). Adenylate kinase then captures the high energy phosphate group present on ADP, resulting in the formation of AMP and adenosine. Adenosine is then able to leave the cell, where it is further degraded to inosine and hypoxanthine (Kloner and Jennings, 2001). 3 These alterations to the metabolic state of the myocyte result in ionic imbalance as well. K+ efflux is facilitated by an increased osmotic load that occurs due to metabolite accumulation (Buja, 2005). The cell will also attempt to correct acidosis via Na+/H+ exchange, which results in increased intracellular Na+ concentrations (Ladilov et al., 1995). Normally, excess intracellular Na+ would be removed by the Na+/K+ ATPase, but ATP depletion prevents this (Kloner and Jennings, 2001). Elevated intracellular Na+ leads to H 2 O influx and cell swelling, as well as the prevention of calcium export via the Na+/Ca2+ exchanger, leading to increased intracellular Ca2+ concentrations. In extreme cases, calcium may be imported by the Na+/Ca2+ exchanger in an attempt to compensate for elevated Na+ levels (Schafer et al., 2001). Intracellular Ca2+ concentration increase also results in activation of proteases, leading to decreased sensitization to Ca2+ in contractile proteins and therefore, impaired myocyte contractility (Buja, 2005). Together, these conditions result in changes to myocardial contractility and a decrease in membrane potential (Kloner and Jennings, 2001), creating an environment that favours the development of ventricular arrhythmia (Buja, 2005). The aforementioned changes are deleterious, but still reversible with timely restoration of oxygen and nutrient supply. However, prolonged/untreated ischemia will eventually result in loss of cell membrane integrity, as the alterations to the metabolic state, combined with elevated intracellular Ca2+ levels activate phospholipases, resulting in the degradation of the phospholipid bilayer. In addition, activated proteases cleave cytoskeletal filaments, which normally anchor the sarcolemma to myofibrils. Eventually, these collective changes result in the compromise of sarcolemmal physical integrity and the death of the myocyte (Buja, 2005). 1.1.3. Reperfusion Injury 4 While prolonged ischemia is obviously deleterious to survival, the restoration of oxygen and nutrient supply - "reperfusion" - is a double-edged sword. During ischemia, ATP is gradually degraded to inosine and hypoxanthine (Kloner and Jennings, 2001), and nitric oxide synthase (NOS) is uncoupled (Raedschelders et al., 2012). Upon reperfusion, four major redox reactions are activated, all resulting in the production of superoxide radicals. Xanthine oxidase converts hypoxanthine to xanthine, and then xanthine to uric acid, yielding a superoxide radical each time. In addition, the uncoupled NOS complex and the activated NAD(P)H oxidase enzyme both oxidize NAD(P)H to NAD(P)+, resulting in the reduction of O 2 to superoxide (Kleikers et al., 2012; Raedschelders et al., 2012). The activation of these redox reactions is exacerbated by a severe increase in arterial flow during the first 5 minutes of reperfusion, resulting in massive reactive oxygen species (ROS) production (Kloner and Jennings, 2001). Finally, reactivation of aerobic respiration results in production of ROS via the electron transport chain (Raedschelders et al., 2012). The increase in free radical presence/production not only serves to deliver oxidative stress damage, but in tandem with elevated intracellular Ca2+ concentrations, triggers mitochondrial permeability transition pore opening (Halestrap et al., 2004). This results in mitochondrial swelling, the collapse of the mitochondrial membrane potential, uncoupling of oxidative phosphorylation, reversal of F 1 F 0 ATPase activity (the promotion of hydrolysis rather than synthesis of ATP), and release of cytochrome c - resulting in the activation of pro-apoptotic caspases (Di Lisa and Bernardi, 2006; Halestrap et al., 2004). Certainly, these deleterious effects are transient (Kloner and Jennings, 2001), but damage caused at the point of reperfusion is often more severe than damage suffered during the ischemic period (but of course, not as severe as the potential damage to a non-reperfused myocardium) (Yellon and Hausenloy, 2007). 5 1.1.4. Post-Reperfusion Inflammation Cascades Myocardial infarction also results in the activation of a myriad of pro-inflammatory cascades. Compromise of cell membrane integrity during MI activates the complement cascade of the innate immune system via release of mitochondrial membrane proteins (Frangogiannis et al., 2002). Complement pathway component mRNA and protein levels were significantly upregulated in infarction zones (Vakeva et al., 1998; Yasojima et al., 1998). As outlined previously, ischemia/reperfusion (I/R) injury provokes the formation of reactive oxygen species. ROS then triggers the release of proteases and damage-associated molecular pattern molecules the latter of which activate Toll-like receptors (TLRs) 2, 4, and 5, eventually resulting in the upregulation of adhesion molecules including vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1), and E-selectin (Carden and Granger, 2000; Marchant et al., 2012; Steffens et al., 2009), as well as pro-inflammatory cytokines such as tumour necrosis factor (TNF)-α, interleukin (IL)-1α, and IL-1β (Ock et al., 2012). Indeed, genetic inactivation of TLR-2 and TLR-4 results in reduced damage following MI (Chong et al., 2004; Shishido et al., 2003). ROS are also capable of acting directly upon endothelial cells, resulting in the release of pro-chemotaxtic factors such as leukotriene B 4 and platelet activating factor (PAF), as well as increased surface expression of P-selectin via triggering release of the adhesion molecule from Weibel-Palade bodies (Carden and Granger, 2000). Indeed, the use of free radical scavengers, such as SOD and catalase, results in reduction of infarction size (Jolly et al., 1984), and genetic overexpression of superoxide dismutase is cardioprotective (Chen et al., 1998; Wang et al., 1998). Finally, I/R injury activates phospholipase A 2 , resulting in increased 6 arachidonic acid levels - which are then subsequently metabolized into pro-inflammatory prostaglandins, eicosanoids, and/or leukotrienes (LTs) (Ahn and Kim, 2012) by circulating neutrophils and/or platelets (Tada et al., 1988). These mechanisms promote the activation and recruitment of peripheral blood leukocytes, resulting in additional sources of pro-inflammatory cytokines present in the proximity of the injury (Carden and Granger, 2000), as well as enhanced leukocyte extravasation into the myocardium (Frangogiannis et al., 2002). Activation of mast cells, neutrophils, and monocytes all promote localized release of pro-inflammatory cytokines and chemokines following ischemia/reperfusion (Frangogiannis et al., 2002; Nahrendorf et al., 2007), as well as upregulation of pro-inflammatory cytokines including IL-1, IL-6, IL-8, monocyte chemotactic protein-1 (MCP-1), and macrophage inflammatory protein-1 (MIP-1) (Frangogiannis et al., 2002; Vandervelde et al., 2007). Leukocyte extravasation first requires the capture of circulating cells by the endothelium. Selectin molecules (P-, L-, and E-selectin) bind to their corresponding ligands, creating leukocyte-endothelial cell connections, and capturing leukocytes close to the endothelium and slowing cell velocity. These cells then roll along the endothelium, allowing CD18-possessing molecules (lymphocyte function-associated antigen (LFA)-1, Mac-1, p150,95) to bind to ICAMs, resulting in firm adhesion of the circulating cell to the endothelial wall, and facilitating transendothelial migration (Kolaczkowska and Kubes, 2013). An increase in the number of adherent leukocytes - especially neutrophils - can cause a phenomenon called "no reflow" where neutrophil accumulation results in capillary plugging following reperfusion (Frangogiannis et al., 2002). Furthermore, adherent neutrophils also release ROS almost immediately upon ICAM1/Mac-1-mediated adhesion, resulting in myocyte injury and death (Albelda et al., 1994; Entman 7 et al., 1992). Studies with animal models have shown that neutrophil depletion (via antiserumtargeting, leukocyte depletion (via whole blood filtration), and inhibition of neutrophil adhesion have all resulted in attenuation of ischemia/reperfusion injury (Marchant et al., 2012). However, leukocyte infiltration is also vital to proper inflammatory response resolution and wound healing. These mechanisms are mainly mediated by monocytes and macrophages. In both mice and humans, monocytes exist in two subsets: pro-inflammatory Ly-6Chi (human analogue: CD14+CD16-) and anti-inflammatory Ly-6Clo (human analogue: CD14lowCD16+) (Nahrendorf et al., 2007; Ziegler-Heitbrock, 2007). Both are involved in wound healing: the proinflammatory subset is recruited by MCP-1, is responsible for debris scavenging, and is present from days 1-4 post-infarction (Kempf et al., 2012; Nahrendorf et al., 2007). Proper clearance of extracellular matrix fragments is vital, as the fragments themselves serve as activators of proinflammatory cells/signaling pathways (Kempf et al., 2012). In contrast, the anti-inflammatory subset is dominant from days 5-10, is recruited by the presence of CX 3 CR1, and mediates proinflammatory signal repression as well as angiogenic factor production (Frangogiannis, 2012; Nahrendorf et al., 2007). The macrophage presence in the myocardium post-infarction is mainly due to monocyte extravasation, which is mediated by C5a, transforming growth factor-β (TGFβ), and MCP-1 (Birdsall et al., 1997; Kumar et al., 1997). The pro-inflammatory environment upregulates macrophage colony-stimulating factor (M-CSF), inducing monocyte differentiation into macrophages (Frangogiannis, 2012). These macrophages then limit the inflammatory response by phagocytosing apoptotic neutrophils and secreting anti-inflammatory mediators such as lipoxin A 4 , TGF-β, and IL-10. Like monocytes, there are two established macrophage subsets: M1 (pro-inflammatory and microbicidal) and M2 (anti-inflammatory and phagocytic), 8 and it is the latter subset that is responsible for inflammatory response resolution post-infarction. Regulatory T lymphocytes are also involved in regulating the inflammatory response, as they secrete IL-10 and TGF-β, in additional to recruiting M2 macrophages to the injured area (Hofmann et al., 2012). The ultimate purpose of these responses is to replace necrotic tissue with granulation tissue, and eventually a collagen-rich scar (Kempf et al., 2012). If the wound healing response is insufficient, scar formation and injury stabilization are impaired, creating a predilection for heart failure (Heymans et al., 1999). However, if the post-infarction inflammatory response is exacerbated, then infarct expansion - the thinning and lengthening of the infarct area post-reperfusion - and adverse ventricular remodeling occurs (Vanhoutte et al., 2007). Thus, a proper balance between pro- and anti-inflammatory mechanisms post-infarction must be maintained in order to limit myocardial injury and preserve heart function. 1.2. Leukotrienes Arachidonic acid is a 20-carbon molecule polyunsaturated fatty acid that serves as the precursor to numerous signaling molecules, termed eicosanoids (from the Greek eikosa, meaning twenty, referring to the number of carbon molecules within their structures). There are four families of eicosanoid molecules: prostaglandins, prostacyclins, thromboxanes, and leukotrienes. The first three are collectively termed “prostanoids,” are formed through the actions of cyclooxygenase (COX) enzymes, and are heavily involved in regulating pain and inflammation (Smyth et al., 2009). On the other hand, leukotrienes, dubbed such because they were first found to be produced by leukocytes and they have three carbon-carbon double bonds in their structures, are formed through the actions of 5-lipoxygenase (5-LO) (Murphy and Gijon, 2007). The 9 leukotriene family consists of five signaling molecules: LTA 4 , B 4 , C 4 , D 4 , and E 4 – the latter three collectively known as the “cysteinyl leukotrienes (cysLTs)” due to a shared common cysteine moiety in their structures (Moos and Funk, 2008). Leukotrienes, although they were not known as such at the time, were first discovered in 1938 by the laboratory of Charles Kellaway in Melbourne, Australia, who identified that treatment with snake venom elicited the secretion of a substance which caused smooth muscle contraction that was not histamine (Feldberg et al., 1938). This substance, termed "slow reacting substance" (SRS), was also found to be released during anaphylaxis (Kellaway and Trethewie, 1940). Two decades later, Brocklehurst identified a “slow reacting substance of anaphylaxis” (SRS-A) that was formed upon antigen stimulus, rather than stored and released like histamine (Brocklehurst, 1960). It would take a further two decades before the identity of SRS-A would be identified. In 1976, Borgeat et al. (1976) discovered that arachidonic acid is oxygenated at the C-5 carbon, and further research revealed a family of metabolites sharing this trait, including what would later be known as LTB 4 (Borgeat and Samuelsson, 1979a) and an unstable intermediate later termed LTA 4 (Borgeat and Samuelsson, 1979b). Structural characterization of SRS-A via highperformance liquid chromatography and mass spectrometry revealed that it was related to the two compounds found previously, and it was also formed from LTA 4 , and thus was termed LTC 4 (Murphy et al., 1979). Identification of leukotrienes D 4 and E 4 would soon follow (Bernstrom and Hammarstrom, 1981; Orning et al., 1980), and all three would be classified as the constituents of SRS-A (Samuelsson, 1983). 1.2.1. Leukotriene Synthesis Pathway 10 The leukotriene synthesis pathway (Figure 1-1) begins with the liberation of arachidonic acid from the perinuclear phospholipid membrane, endoplasmic reticulum, and/or Golgi apparatus by activation of cytosolic phospholipase A 2 α (cPLA 2 α) (Hirabayashi et al., 2004). Arachidonic acid is then converted into LTA 4 via a two-step mechanism through activation of 5-LO. First, arachidonic acid undergoes pro-S hydrogen removal at C-7 with migration of the double bond from C5-C6 to C6-C7, followed by a lipoxygenation reaction at C-5, resulting in the formation of 5-hydroperoxyeicosatetraenoic acid (5-HpETE). Finally, the pro-R hydrogen at C-10 is removed, allowing for the rearrangement of the double bonds and the subsequent migration of the oxygen at C-5 to C-6, forming an epoxide moiety and yielding LTA 4 (Haeggstrom and Funk, 2011). This process is undertaken by 5-LO, but is also facilitated by the action of a second protein, appropriately named 5-lipoxygenase-activating protein (FLAP). Despite being devoid of enzyme activity, FLAP has been shown to stimulate arachidonic acid utilization by 5-LO, as well as 5-LO 5-HpETE-to-LTA 4 conversion efficiency (Abramovitz et al., 1993; Dixon et al., 1990). While the exact mechanism of action is not fully known, research indicates that FLAP acts as a scaffolding protein for 5-LO, and likely presents or transfers arachidonic acid to 5-LO (Poeckel and Funk, 2010). LTA 4 can be converted either into LTB 4 by the enzyme LTA 4 hydrolase (LTA 4 H), or into LTC 4 by LTC 4 synthase (LTC 4 S). LTA 4 H generates a carbocation that delocalizes its charge over the conjugated triene system present from C6-C12 of LTA 4 , leaving the C-12 site open for attack by a H 2 O molecule, which results in the creation of a hydroxyl group. The conjugated triene system shifts to C5-C11, and the epoxide moiety is removed, leaving a hydroxyl group at C5 and 11 Figure 1-1: Leukotriene synthesis pathway. Schematic figure of leukotriene synthesis. Increases in intracellular calcium concentrations promotes the translocation of cytosolic phospholipase A 2 α (cPLA 2 α) and 5-lipoxygenase (5-LO) to the nuclear membrane, where they complex with 5-LO activating protein (FLAP). cPLA 2 α liberates arachidonic acid (AA) from the phospholipid bilayer and presents it to 5-LO, which converts it to leukotriene A 4 (LTA 4 ). LTA 4 is then converted into either leukotriene B 4 (LTB 4 ) by LTA 4 hydrolase (LTA 4 H) present in the cytosol or to leukotriene C 4 by LTC 4 synthase (LTC 4 S), which is also present at the nuclear membrane. LTB 4 and LTC 4 are exported from the cell by multidrug-resistance proteins (MRPs) 4 and 1, respectively. LTB 4 binds to BLT receptors, whereas LTC 4 binds to cysteinyl leukotriene receptors (cysLTRs) 1 and 2. LTC 4 can also be converted to leukotriene D 4 (LTD 4 ), which also binds to CysLT 1 R and CysLT 2 R, by γglutamyl transferase (GGT). Finally, LTD 4 is converted to the stable metabolite leukotriene E 4 (LTE 4 ), which activates the newly discovered CysLT E R, by dipeptidase (DiP). 12 yielding LTB 4 (Haeggstrom and Funk, 2011). Conversely, LTC 4 S activates a glutathione thiol via deprotonation of the sulfhydryl group, the resulting thiolate then attacks LTA 4 at C-6, opening the epoxide ring and conjugating at C-6 to yield LTC 4 . LTC 4 is then converted into LTD 4 by the actions of γ-glutamyl transferase, which removes the glutamate residue from glutathione (Funk, 2001). LTD 4 is in turn converted to LTE 4 by dipeptidase, which removes the glycine residue (Moos and Funk, 2008). LTC 4 S is responsible for the majority of LTC 4 production (Kanaoka et al., 2001), although two related enzymes are also capable of minor LTC 4 production. Microsomal glutathione-S-transferase (mGST) II and mGST III share 44% and 27% homology to LTC 4 S respectively, but where LTC 4 S only conjugates glutathione to LTA 4 , mGST II and mGST III are also able to conjugate glutathione to xenobiotic substances as part of the detoxification process (Penrose, 1999). mGST II is the dominant LTC 4 producing enzyme in liver and endothelial cells (Scoggan et al., 1997; Sjostrom et al., 2001). Synthesis of LTA 4 , LTB 4 , and LTC 4 occur intracellularly. As previously mentioned, increases in intracellular calcium levels promote cPLA 2 α translocation to the perinuclear region, allowing access to phospholipid substrates for the liberation of arachidonic acid (Glover et al., 1995; Schievella et al., 1995). Likewise, 5-LO translocation to the perinuclear membrane is also prompted by elevation of intracellular calcium (Rouzer and Samuelsson, 1985), although there is also evidence that arachidonic acid itself may act on 5-LO to alter the affinity of the latter for the nuclear bilayer (Flamand et al., 2006). 5-LO activity can also be regulated via phosphorylation at Ser271 (MAPKAPK-2 dependent) (Werz et al., 2000), Ser663 (ERK2 dependent) (Werz et al., 2002), and Ser523 (cAMP/PKA dependent) (Luo et al., 2004). Phosphorylation at the first two sites has been shown to activate the enzyme by promoting translocation (Werz et al., 2002; Werz 13 et al., 2000), whereas phosphorylation at Ser523 inactivates 5-LO by preventing translocation (Luo et al., 2005; Luo et al., 2004). Translocation of 5-LO and cPLA 2 α results in the formation of a complex with FLAP, which is already present on the inner nuclear membrane (Christmas et al., 2002). Although most 5-LO expressing cells express 5-LO in the cytoplasm, some cells possess nucleoplasm expression of the enzyme. Migration from cytoplasm to nucleoplasm is mediated by Ser271 phosphorylation (Luo et al., 2003), where subsequent elevations in calcium concentration cause translocation to the inner nuclear membrane, where it can interact directly with FLAP, but not cPLA 2 α or LTC 4 S (Murphy and Gijon, 2007). This phenomenon occurs in neutrophils (Brock et al., 1997), eosinophils (Cowburn et al., 1999), and macrophages (Covin et al., 1998; Woods et al., 1995), and can be triggered by external stimuli or leukocyte-endothelium interactions upon cell adhesion (Cowburn et al., 1999). Furthermore, whether 5-LO is located in the nucleoplasm or in the cytoplasm appears to affect the efficiency of leukotriene production, as research has shown that neutrophils and macrophages with nucleoplasm-localized 5-LO all showed elevated leukotriene production compared to counterparts that possessed cytosolic 5-LO (Brock et al., 1997; Woods et al., 1995). While LTA 4 H is a soluble protein predominantly located in the cytosol that does not translocate, LTC 4 S is found on the outer nuclear membrane and may form heterodimers with FLAP, although they are predominantly positioned on opposite sides of the nuclear membrane (Christmas et al., 2002; Haeggstrom and Funk, 2011). Once synthesized, LTB 4 and LTC 4 are exported from the cell by members of the multidrug resistance-associated protein (MRP) family - specifically MRP4 and MRP1, respectively (Haeggstrom and Funk, 2011). Both of these proteins are members of a group of proteins first identified due to their ability to eliminate amphipathic anions from the cell, thus conferring 14 resistance to pharmacological agents (Kruh and Belinsky, 2003). MRP1 is an ATP-dependent transporter of glutathione and glucuronate conjugates that is located on the basolateral membrane in epithelial cells (Kruh and Belinsky, 2003). Its actions can be blocked via competitive inhibition by the CysLT 1 R pharmacological inhibitor MK571 (Leier et al., 1994), and genetic inactivation of MRP1 results in 25 to 90% higher concentrations of intracellular LTC 4 in tissues that highly express the receptor endogenously (Lorico et al., 1997). MRP4 is also ATPdependent, but transports thiopurine nucleotides and cyclic nucleotides, as well as LTB 4 (Kruh and Belinsky, 2003; Rius et al., 2008). MRP4 requires glutathione as a substrate in order to function as an LTB 4 exporter, and can serve as a LTC 4 exporter in the absence of glutathione (Rius et al., 2008). Interestingly, the expression sites of leukotriene synthesis pathway enzymes do not all coincide, indicating the presence of transcellular synthesis mechanisms for leukotriene production. Whereas cPLA 2 α and LTA 4 H are constitutively expressed in most cells (Haeggstrom and Funk, 2011; Murphy and Gijon, 2007), 5-LO expression is restricted primarily to bone marrow-derived myeloid cells (Radmark and Samuelsson, 2007). Transplantation of 5-LO knockout bone marrow cells into irradiated wild-type mice does not yield any leukotriene production, indicating that bone marrow cells are the ultimate source of all leukotrienes produced in vivo (Zarini et al., 2009). Similarly, LTC 4 S is only expressed in eosinophils, mast cells, monocytes, macrophages (all of which express 5-LO), as well as platelets (which do not express 5-LO) (Lam et al., 1995; Maclouf and Murphy, 1988). Expression of these proteins is upregulated by pro-inflammatory stimuli: cPLA 2 α is upregulated by pro-inflammatory cytokines such as IL-1α, IL-1β, TNF-α (Clark et al., 1995). 5-LO is upregulated by cell differentiation promoting factors such as TGF- 15 β, and granulocyte/macrophage colony-stimulating factor (GM-CSF) (Radmark et al., 2007). 5LO expression is also upregulated upon monocyte transmigration and subsequent differentiation into macrophages (Covin et al., 1998; Pueringer et al., 1992). FLAP expression is also upregulated by GM-CSF stimulation (Pouliot et al., 1994), as well as by pro-inflammatory mediators cytokines such as IL-5 and TNF-α (Cowburn et al., 1999; Reddy et al., 2003), and hypoxia (Gonsalves and Kalra, 2010). Finally, LTC 4 S expression is upregulated by lipopolysaccharide (LPS) stimulation in vivo (Schroder et al., 2005). 1.2.2. Leukotriene Transcellular Biosynthesis As mentioned, 5-LO expression is limited to bone marrow-derived cells (Radmark and Samuelsson, 2007), and while many of these cells also possess either LTA 4 H or LTC 4 S, few cells possess all three of the key enzymes in the leukotriene synthesis cascade. For example, neutrophils, the primary source of LTA 4 generation, do not possess LTC 4 S (Maclouf and Murphy, 1988; Zarini et al., 2009). This indicates that leukotriene biosynthesis involves cooperation between multiple cell types. In vitro experiments have shown that red blood cells are capable of converting exogenously applied LTA 4 into LTB 4 , (Fitzpatrick et al., 1984; Haeggstrom, 2004), whereas platelets (Edenius et al., 1988; Maclouf and Murphy, 1988) and mast cells (Dahinden et al., 1985) can produce LTC 4 under similar circumstances. Similarly, coincubation experiments have shown that LTA 4 produced from neutrophils could be converted to LTB 4 by red blood cells (McGee and Fitzpatrick, 1986) and macrophages (Grimminger et al., 1991), and to LTC 4 by endothelial cells (Brady and Serhan, 1992; Feinmark and Cannon, 1986) and platelets (Maclouf and Murphy, 1988). Neutrophil-produced LTA 4 can also be converted to 16 LTC 4 by keratinocytes (Iversen et al., 1994) and both LTB 4 and LTC 4 by chondrocytes (Amat et al., 1998). Interestingly, different cells possessed different LT production efficiencies – e.g. platelets have been demonstrated to be much more efficient in the production of LTC 4 compared to HUVECs (Folco and Murphy, 2006). Furthermore, LTC 4 production in endothelial cells can be blocked by inactivation of the cell adhesion molecules L-selectin and CD18, indicating that neutrophil adhesion to endothelial cells affects leukotriene production (Brady and Serhan, 1992). These findings have also been confirmed in vivo. Initiation of phagocytosis in whole blood yielded en masse neutrophil production of LTA 4 , and as a result, large amounts of LTB 4 and LTE 4 (likely via platelet production of LTC 4 ) (Fradin et al., 1989). Murine lung tissue has no endogenous sources of LTA 4 (Grimminger et al., 1990), but priming the tissue with endotoxin, followed by stimulation with chemoattractants, resulted in large amounts of LTB 4 and some LTC 4 production, likely produced by the cell clumps consisting of a neutrophil/platelet core surrounded by red blood cells also found in the tissue (Voelkel et al., 1992). In contrast, exogenous application of LTA 4 to the lung resulted in mostly LTC 4 production - likely from pulmonary endothelial cells (Grimminger et al., 1988). In the isolated heart, neutrophil adherence to endothelial cells is instrumental to LTC 4 production (Sala et al., 1996; Sala et al., 1993). In both isolated vessels (Kubes et al., 1991) and animal models of LAD ligation (Sala et al., 2000), inhibition of cell adhesion reduces leukotriene production. LTB 4 and LTD 4 production occurs in the brain post-neutrophil infusion, and was accompanied by an increase in edema (Di Gennaro et al., 2004). 17 Finally, work in knockout and chimeric mouse models has confirmed the existence of leukotriene transcellular biosynthesis. Research has confirmed that neither 5-LO knockout (Funk et al., 1995) nor LTA 4 H knockout mice (Byrum et al., 1999) are capable of LTB 4 production. However, Fabre et al. (2002) demonstrated that when cell populations from both these genotypes are combined, LTB 4 production is possible. Thus, a combination of 5-LO knockout and LTA 4 H knockout leukocytes produced significant levels of LTB 4 upon zymosan stimulation in vitro, whereas the individual populations could not. In addition, transplantation of LTA 4 H knockout bone marrow cells into 5-LO knockout mice resulted in significantly increased ear inflammation and edema in response to arachidonic acid injection (Fabre et al., 2002). Finally, Zarini et al. (2009) demonstrated that transplantation of bone marrow cells from wildtype, LTA 4 H knockout, or LTC 4 S knockout mice into 5-LO knockout mice was capable of restoring production capability for both LTB 4 and cysLTs. 1.2.3. Leukotriene B 4 Leukotriene B 4 exerts its effects via two GPCRs, BLT 1 R and BLT 2 R. BLT 1 R expression was first discovered on phagocytic leukocytes, but later work showed that the receptor was also present on granulocytes, monocytes, dendritic cells, lymphocytes, as well as non-circulating cell types such as vascular smooth muscle cells and endothelial cells. In contrast, BLT 2 R is expressed ubiquitously in humans, with strongest expression levels found in the spleen, liver, ovary, and circulating leukocytes (Back et al., 2011). LTB 4 stimulation exerts extremely potent chemotaxtic effects towards neutrophils (Ford-Hutchinson et al., 1980; Malmsten et al., 1980; Palmblad et al., 1981), eosinophils (Huang et al., 1998; Ng et al., 1991), monocytes (Friedrich et 18 al., 2003; Vaddi and Newton, 1994), and dendritic cells (Shin et al., 2006), as well as lesser effects on basophils (Tanimoto et al., 1992), T lymphocytes (Goodarzi et al., 2003; Leppert et al., 1995), and immature mast cells (Weller et al., 2005). In addition to this, LTB 4 stimulation evokes the release of a host of pro-inflammatory agents from circulating leukocytes, including IL-1β (Rola-Pleszczynski and Lemaire, 1985), IL-6 (Brach et al., 1992; Rola-Pleszczynski and Stankova, 1992), MCP-1 (Huang et al., 2004), while enhancing production of IL-1, IL-2, IL-5, interferon-γ (IFNγ) (Rola-Pleszczynski et al., 1986; Yamaoka and Kolb, 1993), and IL-17 (Chen et al., 2009) in T lymphocytes. Finally, LTB 4 stimulation promotes the upregulation and expression of cell adhesion molecules (Gimbrone et al., 1984; Moraes et al., 2010; Vaddi and Newton, 1994). LTB 4 activation of neutrophils also elicits the release of bactericidal agents and proteinases, including lysozymes (Hafstrom et al., 1981), myeloperoxidase (Terawaki et al., 2005), azurocidin (Di Gennaro et al., 2009), matrix metalloproteinases (MMPs) (Kjeldsen et al., 1992), and α-defensins (Flamand et al., 2007a). The triggering of these responses require greater concentrations of LTB 4 than the chemotaxtic response (Serhan et al., 1982). In addition, LTB 4 stimulation of macrophages increases proliferation (Nieves and Moreno, 2006) and Fc receptormediated phagocytosis (Demitsu et al., 1989; Mancuso and Peters-Golden, 2000) but not efferocytosis (Canetti et al., 2003). Prolonged exposure to LTB 4 also delays apoptosis in neutrophils (Hebert et al., 1996), but not in eosinophils (Murray et al., 2003). Finally, LTB 4 acts on smooth muscle cells to induce vasoconstriction, migration, and proliferation (Back et al., 2004; Sakata et al., 2004). Under basal conditions, LTB 4 has minimal effects on endothelial cells. However, BLT 1 R has been observed in atherosclerotic arteries, indicating that 19 inflammation results in the promotion of LTB 4 -mediated pathways (Back et al., 2004). Indeed, both BLT 1 R (LPS and IL-1β) and BLT 2 R (TNFα) are upregulated by pro-inflammatory stimuli (Qiu et al., 2006b). 1.2.4. Cysteinyl Leukotrienes The cysteinyl leukotrienes, as previously mentioned, consist of LTC 4 , LTD 4 , and LTE 4 . They are produced mainly from eosinophils and mast cells, although endothelial cells, fibroblasts, monocytes and macrophages can also produce cysLTs by transcellular metabolism (Austen, 2005; Carnini et al., 2011; Flamand et al., 2007b; Vannella et al., 2007). Traditionally, LTC 4 and LTD 4 have been considered the active cysLTs, with LTE 4 relegated to an afterthought because of a perception that LTE 4 did not exert biological effects (Austen et al., 2009). However, there is a growing body of literature on LTE 4 -mediated mechanisms, and LTE 4 will receive more attention now that a receptor (GPR99) that binds LTE 4 preferentially has been identified (Kanaoka et al., 2013). Cysteinyl leukotrienes exert their effects locally following secretion - their short half-lives, in tandem with the relative impotency of the stable metabolite LTE 4 , limits their range (Austen et al., 2009) - and can act in a paracrine or autocrine fashion (Carnini et al., 2011; Flamand et al., 2007b; Vannella et al., 2007). Unsurprisingly, given their first identity as SRS-A, cysteinyl leukotrienes are very potent at inducing smooth muscle constriction. CysLT induction of airway smooth muscle constriction is very well studied, and as a result, anti-leukotriene pharmacological agents have become a popular asthma treatment option (Dahlen et al., 1980; Okunishi and Peters-Golden, 2011). 20 CysLTs also affect vascular smooth muscle, and can elicit vasoconstriction in both arteries and veins (Michelassi et al., 1982; Schellenberg and Foster, 1984). Interestingly, cysLTs have been shown to induce vasoconstriction in atherosclerotic coronary arteries, but not in healthy ones (Allen et al., 1998) - indicating that pathogenesis may increase the scale and severity of leukotriene-mediated responses. CysLTs also exert effects on circulating cells and on endothelial cells. CysLT stimulation induces chemotaxis of monocytes (Hashimoto et al., 2009; Ichiyama et al., 2005), eosinophils (Fregonese et al., 2002), dendritic cells (Parameswaran et al., 2004) and CD34+ progenitor cells (Bautz et al., 2001; Boehmler et al., 2009). CysLT stimulation also upregulates Mac-1 in eosinophils (Fregonese et al., 2002), promotes proliferation in CD34+ progenitor cells (Braccioni et al., 2002), and induces release of the chemokine RANTES from platelets (Hasegawa et al., 2010). LTC 4 and LTD 4 stimulation of endothelial cells in vitro results in production of PAF, promotion of neutrophil adhesion (McIntyre et al., 1986), secretion of von Willebrand factor, and surface expression of P-selectin (Datta et al., 1995). In addition, cysLT stimulation triggers vascular hyperpermeability in multiple vascular beds (Dahlen et al., 1981; Hui et al., 2004; Moos et al., 2008). That being said, cysLTs have also been implicated with an important role in host defense, as attenuated leukotriene synthesis has been linked to increased susceptibility to infectious disease in both humans and animal models (Flamand et al., 2007b). While LTE 4 received its fair share of attention following its discovery, its poor affinity for the classical cysLT receptors resulted in its fading from the spotlight. However, early studies noted that LTE 4 potency was 10-fold greater than that of LTC 4 or LTD 4 in guinea pig trachea (Drazen et al., 1982). LTE 4 was also distinct in its ability to augment contractile response to histamine, 21 something that LTC 4 or LTD 4 stimulation could not do (Lee et al., 1984). LTE 4 was also found to induce vascular permeability with equal potency as LTC 4 and LTD 4 when injected intradermally in humans (Soter et al., 1983). These findings suggest that LTE 4 either uniquely mediates pathways that LTC 4 or LTD 4 do not, or that there are receptors (i.e. GPR99) that favour LTE 4 as a ligand over LTC 4 and LTD 4 . 1.3. Cysteinyl Leukotriene Receptors Cysteinyl leukotrienes bind to two G-protein coupled receptors, termed cysteinyl leukotriene receptor 1 (CysLT 1 R) and cysteinyl leukotriene receptor 2 (CysLT 2 R). Further investigation has yielded a receptor favouring leukotriene E 4 rather than C 4 and/or D 4 , recently identified as GPR99 (Kanaoka et al., 2013). In addition to this, the P2Y family of purinergic GPCRs, as well as other GPCRs, has been linked with leukotriene-binding activation. However, the two most well investigated receptors remain the CysLT 1 R and CysLT 2 R. 1.3.1. Cysteinyl Leukotriene Receptor 1 The human CysLT 1 R was first characterized in 1999 by two separate groups. It is a 337 amino acid G-protein coupled receptor that signals predominantly through G q/11 class G proteins, although it does show limited activation of G i/o pathways (Lynch et al., 1999; Sarau et al., 1999). The human CysLT 1 R shares only 38% homology with the human CysLT 2 R, and is actually more similar to the purinoreceptor P2Y 1 (32% homology) than the BLT 1 R (28%) (Heise et al., 2000; Lynch et al., 1999; Sarau et al., 1999). The rank order for ligand affinity was first reported as 22 LTD 4 > LTE 4 = LTC 4 > LTB 4 in transfected Cos-7 cells (Lynch et al., 1999), and LTD 4 > LTC 4 > LTE 4 > LTB 4 in transfected HEK293 cells (Sarau et al., 1999). In humans, CysLT 1 R is most highly expressed in spleen and peripheral blood leukocytes, with lower expression in lung, placenta, small intestine, colon, and thymus (Lynch et al., 1999; Sarau et al., 1999). Later research would pinpoint human peripheral blood leukocyte CysLT 1 R expression to monocytes, macrophages, eosinophils, pre-granulocytic CD34+ cells, B lymphocytes (Figueroa et al., 2001), and mast cells (Mellor et al., 2001). The murine CysLT 1 R was first characterized in 2001 (Maekawa et al., 2001; Martin et al., 2001). It shares ~87% sequence homology with the human CysLT 1 R and exists in two isoforms, a short 337 amino acid variant (consisting of exons I and IV) and a long variant (352 aa total, consisting of exons I, II, III, and IV) that possesses a 13 amino acid extension at the N terminus (Maekawa et al., 2001; Martin et al., 2001). The long isoform is expressed highest in the lung and skin, and more weakly in the heart, kidney, stomach, and tongue. Conversely, the short isoform was found only in lung and skin (Maekawa et al., 2001). Similar to the human receptor, murine CysLT 1 R ligand affinity rank order was reported as LTD 4 > LTE 4 = LTC 4 > LTB 4 (Maekawa et al., 2001; Martin et al., 2001). However, the gap in binding affinity between LTD 4 and LTC 4 is much wider in the mouse, where the EC 50 for LTC 4 is 1,000-fold higher than that of LTD 4 , versus the human, where the difference is only a 10-fold increase (Maekawa et al., 2001). CysLT 1 R is regulated transcriptionally by a myriad of pro- and anti-inflammatory factors – mostly cytokines - depending on the cell type. It is upregulated upon stimulation with IL-4 or IL-13 in human monocytes and monocyte-derived macrophages (Shirasaki et al., 2007; 23 Thivierge et al., 2001; Woszczek et al., 2005), as well as upon IL-5 stimulation in human eosinophils (Thivierge et al., 2000). CysLT 1 R levels in B lymphocytes are upregulated following stimulation by either a combination of IL-4 and activating anti-CD40 antibody, or a combination of IL-4 and the presence of CD154-transfected fibroblasts (Lamoureux et al., 2006). IL-13 treatment results in significant upregulation of both CysLT 1 R mRNA and protein levels, yielding a vulnerability to LTC 4 stimulation-mediated eotaxin production (Chibana et al., 2003). Individual treatment with TGFβ, IFNγ, and IL-13 (but not IL-4) all resulted in CysLT 1 R upregulation in human airway smooth muscle cells (Espinosa et al., 2003). IL-1β induces CysLT 1 R upregulation in human umbilical vein endothelial cells (HUVECs) (Gronert et al., 2001). Conversely, direct IL-10 treatment downregulates CysLT 1 R levels in monocytes (Woszczek et al., 2008), and LPS or zymosan treatment results in CysLT 1 R downregulation via endogenous production of PGE 2 and IL-10 (Thivierge et al., 2006, 2009). Activation of CysLT 1 R results in mobilization of intracellular Ca2+ via both G q/11 and G i/o pathway activation (Baud et al., 1987; Lynch et al., 1999). Confirmed secondary messenger signaling pathways activated include MAPK (Hoshino et al., 1998; Paruchuri et al., 2002), PKC (Hoshino et al., 1998), ERK1/2 (Jiang et al., 2006; McMahon et al., 2000), and PI3K-AktGSK3β (Kim et al., 2010). Induction of proliferation has been observed in numerous cell lines following CysLT 1 R activation, including human hematopoietic cells (Braccioni et al., 2002), epithelial cells (Leikauf et al., 1990; Paruchuri and Sjolander, 2003), smooth muscle cells (Kaetsu et al., 2007; Panettieri et al., 1998), and astrocytes (Fang et al., 2006; Huang et al., 2008). In addition, CysLT 1 R activation upregulates pro-inflammatory mediators, including βintegrins (Boehmler et al., 2009; Massoumi and Sjolander, 2001), IL-4 (Bandeira-Melo et al., 24 2002), IL-5 (Faith et al., 2008; Frieri et al., 2003; Hasegawa et al., 2010), IL-8 (Thompson et al., 2006), IL-11 (Lee et al., 2007), TGF-β1 (Kato et al., 2005), as well as MIP-1α and MIP-1β (Ichiyama et al., 2009). Finally, CysLT 1 R pharmacological blockade results in decreased levels of IL-4, IL-5, IL-8, IL-11, IL-13, TNFα, RANTES, and TGF-β (Henderson et al., 2002; Kiwamoto et al., 2011; Lee et al., 2007; Ueda et al., 2003). 1.3.2. Cysteinyl Leukotriene Receptor 2 The human CysLT 2 R was first characterized in 2000 by three independent groups (Heise et al., 2000; Nothacker et al., 2000; Takasaki et al., 2000). It is a 346 aa protein that shares 33% homology with the orphan receptor GPR17 (and as mentioned before, only 38% homology with CysLT 1 R) (Heise et al., 2000). CysLT 2 R binds LTD 4 and LTC 4 with equal affinity and signals through G q/11 (Heise et al., 2000). LTE 4 behaves as a partial agonist, eliciting a calcium response in transfected HEK293 cells (Nothacker et al., 2000; Takasaki et al., 2000) and Cos-7 cells (Heise et al., 2000), but LTB 4 does not (Heise et al., 2000; Takasaki et al., 2000). Furthermore, these responses are unaffected by CysLT 1 R pharmacological antagonism (Heise et al., 2000; Nothacker et al., 2000; Takasaki et al., 2000). The human CysLT 2 R expression profile is distinct from that of CysLT 1 R, being found predominantly in the heart (expressed in atria, ventricles, and Purkinje fibres - but not the aorta), spleen, brain, lymphatic system, placenta, and adrenal gland (Heise et al., 2000; Nothacker et al., 2000; Takasaki et al., 2000). Indeed, it is the dominant cysLT receptor in heart and brain (Takasaki et al., 2000), and unlike CysLT 1 R, is not found in high abundance in the lung (Heise et al., 2000). CysLT 2 R is also expressed by numerous circulating cells, including eosinophils (Mita et al., 2001), monocytes, macrophages (Heise et al., 25 2000), mast cells (Mellor et al., 2003) and platelets (Hasegawa et al., 2010), but not neutrophils (Figueroa et al., 2003). It is also expressed in HUVECs (Lotzer et al., 2003) and coronary artery SMCs (Kamohara et al., 2001). The murine CysLT 2 R was characterized in 2001 (Hui et al., 2001) and shares 74% homology with its human counterpart (Evans, 2003). It is a 309 aa protein primarily expressed in spleen, thymus, and adrenal gland, with weaker expression in kidney, brain, and peripheral blood leukocytes. While Northern blot analysis failed to detect CysLT 2 R in the murine heart, the more sensitive in situ hybridization technique was able to detect CysLT 2 R in murine Purkinje cells and cardiac endothelial cells (Hui et al., 2001). Apart from this, however, further elucidation of specific cell types expressing CysLT 2 R in the mouse has been elusive due to lack of a specific antibody for the receptor. Using a mouse model where LacZ was placed under the Cysltr2 promoter, Moos et al. (2008) was able to detect CysLT 2 R expression in cardiac tissue (Jiang et al., 2008), as well as microvasculature in brain, bladder, skin, ear, and cremaster muscle. Further studies demonstrated CysLT 2 R expression in murine neurons of the myenteric and submucosal plexus in the small intestine, colonic myenteric plexus, dorsal root ganglia, and nodose ganglion (Barajas-Espinosa et al., 2011), as well as retinal microvasculature and resident pericytes (Barajas-Espinosa et al., 2012). CysLT 2 R expression is upregulated by pro-inflammatory cytokines, including IFNγ (Woszczek et al., 2007), IL-4 (Lotzer et al., 2003), IL-13 (Shirasaki et al., 2007), and IL-18 (Zhou et al., 2009). Pretreatment with IFNγ enhances responsiveness to cysLT stimulation in endothelial cells (Woszczek et al., 2007), monocytes, T cells, and B lymphocytes (Early et al., 2007). 26 CysLT 2 R is also upregulated in murine models following ischemia in the heart (Jiang et al., 2008), retina (Barajas-Espinosa et al., 2012), and brain (Shi et al., 2012b). Interestingly, CysLT 2 R expression is suppressed by TNFα, LPS, or IL-1β (Sjostrom et al., 2003) - possibly as a mechanism designed to limit the extent of inflammation. CysLT 2 R activation results in altered endothelial cell function, as well as cytokine secretion. Activation of CysLT 2 R in HUVECs leads to increased intracellular calcium concentration, myosin light chain kinase activation (Carnini et al., 2011; Lotzer et al., 2003), P-selectin surface expression (Pedersen et al., 1997), and the upregulation of a myriad of pro-inflammatory genes including CXCL2 (which encodes the MIP-2α protein), SELE (E-selectin), IL-8, EGR1, and PTGS2 (Uzonyi et al., 2006). In human mast cells, CysLT 2 R activation facilitates IL-8 secretion, but through a pertussis toxin-sensitive pathway (Mellor et al., 2003). In animal models, a murine dextran sulfate sodium-induced colitis model showed attenuated TNFα secretion in the absence of functional CysLT 2 R (Barajas-Espinosa et al., 2011), and a murine model of endothelium-targeted transgenic CysLT 2 R upregulation presented vascular hyperpermeability in response to stimulation by both leukotrienes or IgE (Hui et al., 2004; Moos et al., 2008). This mechanism appears to be regulated, at least partially, by Ca2+ signaling and transendothelial vesicle transport (Moos et al., 2008). 1.3.3. Other Cysteinyl Leukotriene Receptors In addition to CysLT 1 R and CysLT 2 R, data has implicated cysLT activation of other GPCRs. A possible third cysLT receptor was first proposed after Mellor et al. (2001) noted that IL-4 27 treatment in mast cells upregulated two receptors: CysLT 1 R and a receptor that could not be CysLT 2 R based on pharmacological antagonism experiments. Moreover, these cells responded to both cysLTs and UDP, and as CysLT 1 R activation by UDP was later ruled out (Capra et al., 2005; Woszczek et al., 2010), this indicated that the unknown receptor had purinergic signaling capability. Further investigation revealed that GPR17, a receptor structrually and phylogenetically related to other P2Y receptors and cysLT receptors (Daniele et al., 2011), could be activated in a specific and dose-dependent manner by both cysLTs and uracil nucleotides (Ciana et al., 2006; Daniele et al., 2011). In addition, GPR17 heterodimerizes with CysLT 1 R and acts as a negative regulator of CysLT 1 R without activating any responses of its own or the need for uracil nucleotide stimulation (Maekawa et al., 2009; Maekawa et al., 2010). However, GPR17 knockdown abolished LTD 4 -induced effects on cell viability (Daniele et al., 2010). Thus, the question was: does GPR17 respond directly to cysLT stimulation or act on cysLT receptors upon activation by uracil nucleotides (Back et al., 2011; Benned-Jensen and Rosenkilde, 2010)? The answer appears to be both: GPR17 has two binding sites - one for cysLTs and the other for nucleotides (Parravicini et al., 2010; Parravicini et al., 2008). Receptor activation via either cysLTs or nucleotides induces a change in conformational state that enhances functional responsiveness to both sets of agonists. In addition, both sets of agonists trigger homologous desensitization, but interestingly, while UDP-glucose stimulation desensitizes GPR17 to LTD 4 -mediated responses, LTD 4 stimulation does not reciprocate, and has no effect on sensitivity to UDP-glucose-mediated responses (Daniele et al., 2011). LTE 4 has long been regarded as a biological marker for the production of its more potent siblings LTC 4 and LTD 4 . However, the stability of LTE 4 also ensures that it is the most 28 abundant cysLT in vivo, and it has been shown to induce smooth muscle contraction and eosinophilia (Gauvreau et al., 2001; Lee et al., 1984; Lewis et al., 1980). Paruchuri et al. (2008) reported that LTE 4 mediated prostaglandin D 2 production in human mast cells. Curiously, while this response was attenuated by the CysLT 1 R antagonist MK-571, it was not affected by CysLT 1 R genetic knockdown alone, nor could it be triggered by LTD 4 stimulation, suggesting the existence of a novel LTE 4 -selective receptor. Later work would reveal that this response was mediated by the purinergic receptor P2Y 12 (Paruchuri et al., 2009). Indeed, Nonaka et al. (2005), using a computer model, determined that LTE 4 is a potential ligand for P2Y 12 . However, Foster et al. (2013) reported no signs of P2Y 12 activation in transfected HEK-293 cells. Furthermore, they saw no response in extracted platelets, ruling out that P2Y 12 activation exerts its effects through heterodimerization with another receptor. Around the same time as Paruchuri et al., Maekawa et al. (2008) reported that CysLT 1 R/CysLT 2 R double knockout mice still showed ear vascular leakage post-cysLT stimulation, and that LTE 4 elicited a much more robust response than LTC 4 or LTD 4 in these mice. However, the LTE 4 -mediated response reported by Maekawa et al. was only sensitive to the CysLT 1 R-selective antagonist MK-571 in wild-type mice, and no inhibition was noted when CysLT 1 R knockout mice were treated with MK-571 (Maekawa et al., 2008). Furthermore, CysLT 2 R knockout mice treated with MK-571 actually showed elevated permeability relative to wild-type and CysLT 1 R knockout mice, indicating that this putative receptor - termed CysLT E R at the time - may be negatively regulated by the classical cysLT receptors (Maekawa et al., 2008). This receptor has been recently identified as GPR99 (Kanaoka et al., 2013), a receptor previously believed to be a purinergic receptor before research revealed that it is actually 29 activated by α-ketoglutarate (Abbracchio et al., 2005; He et al., 2004; Qi et al., 2004). Indeed, the LTE 4 -mediated ear edema observed in CysLT 1 R/CysLT 2 R double knockout mice was almost completely abolished in GPR99/CysLT 1 R/CysLT 2 R triple knockout mice. Stimulation with LTC 4 or LTD 4 did elicit a mild response in CysLT 1 R/CysLT 2 R double knockout mice, indicating that GPR99 can bind LTC 4 or LTD 4 in the absence of the classical cysLT receptors (Kanaoka et al., 2013). 1.4. Leukotriene Involvement in Pathology Given their potent pro-inflammatory effects, it is not surprising that leukotrienes have been linked with pathogenesis for numerous conditions, including inflammatory bowel disease, cholestasis, hepatic inflammation, arthritis, and atopic dermatitis (Back et al., 2011; Haeggstrom and Funk, 2011). However, the conditions that have received the most attention are: asthma and other inflammatory diseases of the respiratory tract, cancers, and ischemic injury in the cerebroand cardiovascular systems. 1.4.1. Asthma/Respiratory Diseases Leukotrienes were first discovered as mediators of bronchial smooth muscle constriction, so it is understandable that they are involved in asthma, as well as other pathologies involving inflammation and/or constriction of the airways. Elevated levels of cysLTs have been measured in bronchoalveolar lavage (BAL) fluid, urine, and blood from asthmatic adults, and cysLTs are elevated in BAL fluid taken at night from nocturnal asthmatics (Taylor et al., 1992; Wenzel et 30 al., 1990; Wenzel et al., 1995). LTB 4 also plays a role in asthma: it is elevated in asthmatic patients (Wenzel et al., 1995) and murine studies show that BLT 1 R plays a vital role in mediating Th-2 cytokine mediated airway inflammation (Miyahara et al., 2005), whereas BLT 1 R-expressing T cells are upregulated in allergic asthma patient airways (Islam et al., 2006). Despite its lack of affinity for the CysLT 1 R and CysLT 2 R, LTE 4 also induces bronchial contraction with equal potency as LTC 4 and LTD 4 (Davidson et al., 1987), a finding that makes more sense now following the revelation of the CysLT E R (GPR99). Also interesting was that asthmatic airways maintained sensitivity to LTE 4 , whereas responses to LTC 4 and LTD 4 were attenuated compared to equivalent responses in healthy individuals (Adelroth et al., 1986; O'Hickey et al., 1988; Weiss et al., 1982). LTE 4 , but not LTC 4 and LTD 4 , was also able to induce hyperresponsiveness to histamine (Lee et al., 1984). Now that a receptor preferential for LTE 4 has been identified (Kanaoka et al., 2013; Maekawa et al., 2008), the role of CysLT E R activation in asthma and bronchoconstriction warrants further attention. As of 2011, there were three CysLT 1 R antagonists and one 5-LO antagonist clinically available (Okunishi and Peters-Golden, 2011). The lukast family of CysLT 1 R antagonists, all first discovered in the 1990s, consists of pranlukast (Obata et al., 1992), zafirlukast (Krell et al., 1990), and montelukast (Jones et al., 1995). Of the three, montelukast is the most widely available and promoted, whereas pranlukast is only available in Japan and other Asian countries (Okunishi and Peters-Golden, 2011). There are numerous studies detailing the ability of CysLT 1 R antagonism to mitigate inflammatory bronchospastic responses in cases of acute asthma (Dockhorn et al., 2000; Gaddy et al., 1992), persistent asthma (Reiss et al., 1998; Suissa et al., 1997), aspirin-triggered asthma (Christie et al., 1991; Fischer et al., 1994), and in response 31 to allergens (Taylor et al., 1991). CysLT 1 R blockade also resulted in a marked decreased in airway histamine and TNF-α levels (Figueroa et al., 2001). However, CysLT 1 R antagonism failed to abrogate eosinophil recruitment (Diamant et al., 1999). The 5-LO inhibitor zileuton was discovered in 1991 (Carter et al., 1991), and was considered to have more potential than cysLT receptor antagonists given its ability to block both cysLT receptor and BLT receptor signaling. While it has similar anti-bronchoconstriction properties to the CysLT 1 R antagonists (Israel et al., 1996; Liu et al., 1996; Wenzel et al., 1995), it has not gained widespread use because of an initial need for four daily doses and the possibility of hepatocellular injury (Okunishi and Peters-Golden, 2011). Furthermore, there is little evidence to suggest that zileuton is superior to any of the lukast family members (Israel et al., 1996; Wenzel et al., 2007). While these compounds do improve lung and airway function while decreasing disease symptoms, they are still not as efficacious as inhaled corticosteroids in the treatment of asthma (National Asthma and Prevention, 2007; Wenzel, 2003). A further complication is that there is a distinct and significant subset of patients who are unresponsive to leukotriene pathway modifiers (Barnes et al., 2005; Terashima et al., 2002). Despite this, these compounds are effective in treating other conditions such as aspirin-induced asthma, exercise-induced bronchoconstriction (Dahlen, 2006), allergic rhinitis, and chronic obstructive pulmonary disease (Back et al., 2011). They also provide a tested alternative therapy option for individuals unwilling or incapable of using inhaler devices or inhaled corticosteroids (Okunishi and Peters-Golden, 2011). 1.4.2. Cancer 32 CysLT signaling activates secondary messengers that have been implicated in cell survival and proliferation. LTD 4 stimulation upregulates COX-2, via activation of ERK1/2, which in turn activates Bcl-2 signaling, promoting cell survival (Wikstrom et al., 2003). Pharmacological blockade of COX-2 resulted in increased apoptosis rate, as well as increased pro-apoptotic pathway signaling. This effect in turn is countered by additional LTD 4 stimulation (Ohd et al., 2000). Research has also shown that LTC 4 stimulation induces endothelial cell proliferation (Modat et al., 1987), whereas LTD 4 stimulation encourages hematopoietic progenitor cell proliferation (Boehmler et al., 2009; Braccioni et al., 2002). In addition, LTB 4 levels are elevated in human colon (Larre et al., 2008) and pancreatic (Ding et al., 2005) cancers, and LTB 4 stimulation promotes proliferation in colon cancer cell lines (Ihara et al., 2007), pancreatic cancer cell lines (Tong et al., 2005), as well as in neural (Wada et al., 2006) and hematopoietic stem cells (Chung et al., 2005). LTB 4 , LTC 4 , and LTD 4 stimulation are pro-angiogenic (Kim et al., 2009; Tsopanoglou et al., 1994). Inhibition of leukotriene synthesis pathway components has been implicated to impede cancer progression in some pre-clinical models. Pharmacological inhibition of 5-LO prevents tumorigenesis in murine lung following treatment with the carcinogen nicotine-derived nitrosamine ketone (Rioux and Castonguay, 1998), and COX-2 and 5-LO blockers in tandem had additive effects in blocking tumour growth in mice xenografted with human colon (Ye et al., 2005), esophageal (Chen et al., 2004b), breast (Barry et al., 2009), and skin (Fegn and Wang, 2009) cancer cells. Blockade of LTA 4 H has been shown to reduce tumour load in rat esophageal adenocarcinomas (Chen et al., 2003), and LTA 4 H downregulation inhibited in vitro and in vivo colon cancer cell growth (Jeong et al., 2009). 33 Both BLT receptors (Hennig et al., 2005) and cysLT receptors (Magnusson et al., 2007; Matsuyama et al., 2007; Ohd et al., 2003) have been found in abundance in cancerous cells and tissues. Appropriately, BLT receptor blockade has been shown to induce apoptosis in colon cancer cells (Ihara et al., 2007) and inhibit tumour growth and metastasis in murine pancreas following implantation with pancreatic cancer cells (Hennig et al., 2005). There is an interesting dichotomy regarding the cysteinyl leukotriene receptors where CysLT 1 R expression appears to be deleterious and CysLT 2 R expression beneficial. Indeed, high CysLT 1 R combined with low CysLT 2 R expression has been associated with increased malignancy and reduced survival. Conversely, low CysLT 1 R combined with high CysLT 2 R expression has been linked with increased survival (Magnusson et al., 2007; Magnusson et al., 2011; Matsuyama et al., 2007). CysLT 1 R signaling promotes cell proliferation via activation of the ERK1/2 signaling pathway (Paruchuri and Sjolander, 2003), and receptor inhibition reduces tumour burden in a mouse model of lung cancer (Gunning et al., 2002), and induces apoptosis in prostate cancer cell lines (Matsuyama et al., 2007). Less is known about the role of CysLT 2 R in tumorigenesis, but there is evidence that it is involved in promotion of apoptosis and terminal differentiation in colon caricinoma cell lines (Magnusson et al., 2007). 1.4.3. Ischemic/Cardiovascular Disease Given the depth and breadth of leukotriene involvement in inflammatory responses, it was only natural that their potential involvement in cardiovascular disease would be investigated. Indeed, urinary excretion of LTE 4 is elevated following myocardial infarction, indicating leukotriene production during the event (Allen et al., 1993; Carry et al., 1992). Murine models also show a 34 similar phenomenon after LAD ligation (Sala et al., 2000). Furthermore, the enzymes involved in leukotriene production have been found in atherosclerotic plaques, and human coronary arteries express 5-LO, FLAP, and LTA 4 H and respond to LTC 4 and LTD 4 (Allen et al., 1998). In addition, 5-LO, FLAP and LTC 4 S are all upregulated in aneurysm thrombi. Gene variants within the leukotriene synthesis pathway have been associated with increased atherosclerosis, myocardial infarction, and stroke (Haeggstrom and Funk, 2011). Dwyer et al. (2004) examined a population of Angelenos and found that individuals possessing variant alleles for ALOX5, the gene encoding 5-LO, had increased intima-media thickness and elevated plasma levels of the inflammatory marker C-reactive protein. A study of the Icelandic population then revealed that polymorphisms in ALOX5AP, the gene encoding FLAP, conferred a greater risk of myocardial infarction and stroke (Helgadottir et al., 2004) - these findings were then replicated in a Scottish population (Helgadottir et al., 2005). However, further research has yielded contradictory results, with some studies confirming that ALOX5AP polymorphisms were associated with myocardial infarction (Linsel-Nitschke et al., 2008), coronary heart disease, (Huang et al., 2010) and stroke (Lee et al., 2011; Sharma et al., 2013). Conversely, other studies have failed to identify any association between ALOX5AP polymorphisms and stroke risk (Barral et al., 2012; Kostulas et al., 2007; Lemaitre et al., 2009; Zintzaras et al., 2009). LTA4H gene variants have also been linked with increased myocardial infarction risk (Helgadottir et al., 2006), whereas LTC4S gene variants have been linked with stroke, but not ischemic heart disease or atherosclerosis in a Danish population (Freiberg et al., 2009). This association with stroke was also found in British and Chinese studies (Bevan et al., 2008; Wang et al., 2012). 35 Early studies investigating the role of cysteinyl leukotrienes in CVD were largely optimistic. Pharmacological inhibition of 5-LO or use of a lipoxygenase/cycloxygenase dual antagonist resulted in decreased infarction size (Amsterdam et al., 1993; Hashimoto et al., 1990; Ito et al., 1989). Use of the non-selective peptidoleukotriene receptor antagonist LY171883 or LY203647 yielded mixed results - whereas the former attenuated infarction size following ischemia (Hock et al., 1992), the latter failed to have any effect on infarction damage (Hahn et al., 1992). Administration of the CysLT 1 R antagonist pranlukast resulted in significantly reduced infarction size (Ito et al., 1989; Toki et al., 1988). These studies targeting the cysLTs did not report reduction in leukocyte recruitment and extravasation following cysLT receptor antagonism (Hock et al., 1992; Ito et al., 1989; Toki et al., 1988), although 5-LO antagonism, but not lipoxygenase/cycloxygenase dual antagonism (Amsterdam et al., 1993), was reported to decrease neutrophil accumulation in the infarcted tissue (Ito et al., 1989; Toki et al., 1988). While most of these studies reported a beneficial effect following pharmacological antagonism of the leukotriene synthesis pathway, indicating a clear role for leukotrienes in myocardial infarction damage, they are somewhat contradictory regarding the mechanisms/actions affected by drug treatment. Carnini (2011) and Back (2011) have both attributed this to the non-selective nature of these early pharmacological agents - particularly those targeting the cysLT receptors - and also posit that the early compounds used may have partial agonist activity or may have not displayed sufficient potency to compete with elevated amounts of local endogenous ligands at the site of injury. Indeed, later studies using montelukast have shown that CysLT 1 R antagonism possesses antiatherogenic effects (Ge et al., 2009; Jawien et al., 2008; Kaetsu et al., 2007). While both CysLT 1 R and CysLT 2 R are expressed in diseased human arteries (Allen et al., 1998; Spanbroek et al., 2003), CysLT 2 R is the dominant isoform in the myocardium (Heise et al., 36 2000) - but the lack of a selective and potent pharmacological agent for the receptor until 2010 (Wunder et al., 2010) has made identifying CysLT 2 R-specific effects difficult. CysLT 2 R is also upregulated in the heart following ischemia (a finding that is corroborated in other organs such as the retina and brain (Barajas-Espinosa et al., 2012; Jiang et al., 2008; Shi et al., 2012b). Hui et al. (2004) generated a transgenic mouse model where seven copies of the human CYSLTR2 gene under control of the murine Tie2 promoter in order to create an “endothelial-targeted” CysLT 2 R overexpressing model. These mice showed exacerbated vascular permeability in response to cysLT stimulation in ear, retina, and cremaster microvasculatures (Barajas-Espinosa et al., 2012; Hui et al., 2004; Moos et al., 2008), significantly increased infarction damage (that was abrogated using the non-selective CysLT receptor antagonist Bay-u9773) accompanied by increased edema, leukocyte extravasation, and elevated expression of adhesion molecules (Jiang et al., 2008). Also, the CysLT 2 R overexpressing mouse model shows accelerated left ventricular remodeling at two weeks post-infarction (Jiang et al., 2008). Ischemic disease, predominantly stroke, occurs in the brain as a result of thrombosis or hemorrhage. Arachidonic acid is released due to degradation of membrane lipids during ischemia, and this release is greatest in brain regions most susceptible to ischemia/reperfusion injury (Katsuki and Okuda, 1995; Westerberg et al., 1987). Indeed, leukotriene production has been noted in the brain during ischemia, and dramatically increased following reperfusion (Moskowitz et al., 1984; Rao et al., 1999). Both CysLT 1 R and CysLT 2 R are expressed in the brain (Heise et al., 2000; Lynch et al., 1999), and both are upregulated following ischemia (Fang et al., 2007; Shi et al., 2012b; Zhao et al., 2011). CysLT 1 R in the brain is primarily localized to microvascular endothelial cells, with traumatic injury inducing expression in neurons and glial 37 cells (Zhang et al., 2004). Treatment with CysLT 1 R-selective antagonists results in decreased ischemic damage and improved neurological function (Fang et al., 2006; Zhang et al., 2002). In addition, pranlukast treatment also decreased brain cryoinjury extent in mice (Ding et al., 2007), indicating a possible role for leukotrienes in traumatic brain injury as well. CysLT 2 R is highly expressed in the central nervous system, including the hypothalamus, thalamus, putamen, pituitary, and medulla (Heise et al., 2000). In addition, the receptor is also found in the brain microvasculature (Moos et al., 2008). Di Gennaro et al. (2004) have found that neutrophils are the source of cysLT synthesis in the brain, and that neutrophil activation results in a brain-blood barrier disruption that can be prevented with FLAP pharmacological antagonism. In addition, exogenous addition of LTB 4 did not elicit similar effects as neutrophil activation, and Bay-u9773 was more effective than montelukast in preventing blood-brain barrier disruption - indicating the involvement of CysLT 2 R. In vitro induction of ischemia in medulla cells resulted in increased cell death in cells overexpressing CysLT 2 R. Treatment of these cells with LTD 4 induced apoptosis, which was inhibited by Bay-u9773 but not montelukast (Sheng et al., 2006). In vivo, use of the CysLT 2 R-selective antagonist HAMI3379 resulted in reduced infarct volume, edema, and neuronal degredation and neuronal death (Shi et al., 2012a). CysLT stimulation has also been shown to induce astrocyte proliferation at low concentrations through a CysLT 1 R-mediated process and astrocyte death at high concentrations through a CysLT 2 R-mediated process (Huang et al., 2008). In addition, ischemia resulted in upregulation of CysLT 2 R in astrocytes while inducing cell death via CysLT 2 R-mediated aquaporin 4-upregulation (Qi et al., 2011). Ischemia results in the acute phase (6-24 hours post-ischemia) upregulation of CysLT 2 R in injured neurons within the infarction zone, and the late phase (3-28 days) upregulation of CysLT 2 R in 38 microglia and astrocytes, indicating a role for CysLT 2 R in astrogliosis and microgliosis postischemia (Zhao et al., 2011). 1.5. Research Objectives and Hypotheses Although endothelium-targeted CysLT 2 R overexpression has been linked with increased myocardial infarction damage and vascular hyperpermeability, the precise mechanisms underlying these phenomena remain unresolved. Investigation of the role of CysLT 2 R activation in myocardial infarction injury has been difficult due to the absence of receptor-selective pharmacological antagonists until very recently. Our aims are thus fourfold: 1) use these novel pharmacological compounds to examine if CysLT 2 R is the main mediator of the exacerbated myocardial infarction damage phenotype observed in CysLT 2 R-overexpressing mice; 2) determine whether CysLT 2 R activation is involved during ischemia, at reperfusion, or during the inflammatory response following infarction; 3) examine whether the difference in injury severity in CysLT 2 R-overexpressing mice post-acute ischemia translates into altered cardiac function long-term; and 4) examine the molecular mechanisms underlying CysLT 2 R regulation of vascular permeability and leukocyte behaviour. We hypothesize that the exacerbated injury phenotype observed in CysLT 2 R-overexpressing mice is primarily mediated by CysLT 2 R (rather than other receptors), and that CysLT 2 R activation most strongly impacts the inflammatory response post-reperfusion. In addition, we believe that exacerbation of CysLT 2 R-mediated myocardial damage post-infarction will result in decreased long term cardiac function and 39 pathological cardiac remodeling. Lastly, CysLT 2 R activation likely mediates vascular permeability via localized leukotriene production and promotion of leukocyte recruitment to the affected site. By investigating and understanding how CysLT 2 R activation results in exacerbation of myocardial infarction injury, we hope to establish that CysLT 2 R is a viable therapeutic target for the treatment of cardiovascular disease. 40 Chapter 2: A selective cysteinyl leukotriene receptor 2 antagonist blocks myocardial ischemia/reperfusion injury and vascular permeability in mice 41 2.1. Abstract Cysteinyl leukotrienes (cysLTs) are potent inflammatory mediators that predominantly exert their effects by binding to cysteinyl leukotriene receptors of the G protein-coupled receptor family. CysLT receptor 2 (CysLT 2 R), expressed in endothelial cells of some vascular beds, has been implicated in a variety of cardiovascular functions. Endothelium-specific overexpression of human CysLT 2 R in transgenic mice (EC) greatly increases myocardial infarction damage. Investigation of this receptor, however, has been hindered by the lack of selective pharmacological antagonists. Here, we describe the characterization of 3-(((3carboxycyclohexyl)amino)carbonyl)-4-(3-(4-(4-phenoxybutoxy)phenyl)-propoxy)benzoic acid (BayCysLT 2 ) and explore the selective effects of this compound in attenuating myocardial ischemia/reperfusion damage and vascular leakage. Using a recently developed β-galactosidaseβ-arrestin complementation assay for CysLT 2 R activity (Yan et al., 2011), we determined BayCysLT 2 to be ~20-fold more potent than the nonselective dual cysLT receptor 1 (CysLT 1 R)/CysLT 2 R antagonist 4-(((1R,2E,4E,6Z,9Z)-1-((1S)-4-carboxy-1-hydroxybutyl)2,4,6,9-pentadecatetraen-1-yl)thio)benzoic acid (Bay-u9773) (IC 50 274 nM versus 4.6 μM, respectively). Intracellular calcium mobilization in response to cysteinyl leukotriene administration showed that BayCysLT 2 was >500-fold more selective for CysLT 2 R compared with CysLT 1 R. Intraperitoneal injection of BayCysLT 2 in mice significantly attenuated leukotriene D 4 -induced Evans blue dye leakage in the murine ear vasculature. BayCysLT 2 administration either before or after ischemia/reperfusion attenuated the aforementioned increased myocardial infarction damage in EC mice. Finally, decreased neutrophil infiltration and leukocyte adhesion molecule mRNA expression were observed in mice treated with 42 antagonist compared with untreated controls. In conclusion, we present the characterization of a potent and selective antagonist for CysLT 2 R that is useful for discerning biological activities of this receptor. 43 2.2. Introduction Myocardial infarction is the leading killer of both males and females in the industrialized world (Yellon and Hausenloy, 2007). The condition is characterized by a cessation of oxygen/nutrient supply to the myocardium, usually caused by a blockage of the coronary circulation. Although the only remedy to this problem is timely restoration of blood flow (“reperfusion”), this results in the en masse formation of reactive oxygen species, creating oxidative stress (Zweier, 1988). Reactive oxygen species formation, in tandem with chemoattractant release during myocardial infarction, invokes the innate inflammatory response, resulting in adhesion protein up-regulation, leukocyte infiltration, and extracellular matrix degradation (Vinten-Johansen, 2004). It is these post-reperfusion events that contribute the majority of myocardial infarction-induced damage (Yellon and Hausenloy, 2007). The cysteinyl leukotrienes (cysLTs), so termed because of the common cysteine moiety in their structures, are a group of proinflammatory arachidonic acid metabolites consisting of LTC 4 , LTD 4 , and LTE 4 . CysLTs exert their effects primarily by binding two characterized G proteincoupled receptors: cysLT receptor 1 (CysLT 1 R) (Lynch et al., 1999) and cysLT receptor 2 (CysLT 2 R) (Heise et al., 2000). CysLTs have also been shown to interact with the putative cysteinyl leukotriene E receptor (Maekawa et al., 2008), the P2Y 12 receptor (purinergic receptor P2Y, G protein-coupled, 12) (Nonaka et al., 2005), and G protein-coupled receptor 17 (Ciana et al., 2006; Maekawa et al., 2009). Of these receptors, CysLT 1 R is the most extensively studied (Moos and Funk, 2008) and has been shown to mediate bronchoconstriction and airway inflammation (Funk, 2005). CysLT 2 R has not received as much attention as CysLT 1 R, although 44 there has been a wave of interest concerning this receptor in cardiovascular disease (Funk, 2005). CysLT 2 R is expressed in multiple organs in the mouse, including the heart (Jiang et al., 2008), brain, cremaster muscle vasculature (Moos et al., 2008), and myenteric neurons (BarajasEspinosa et al., 2011). In humans, it is found in umbilical cord endothelial cells (Lotzer et al., 2003; Uzonyi et al., 2006), myocardium, and coronary vessels (Kamohara et al., 2001). CysLT 2 R mediates vascular endothelium permeability via a vesicular transcytotic mechanism (Hui et al., 2004; Moos et al., 2008) and is regulated by transcriptional activation of chemokine secretion (Thompson et al., 2008). The role of enzymes involved in leukotriene biosynthesis and the cysLT receptors in the cardiovascular context is being studied extensively. Research has shown that single-nucleotide polymorphisms in the genes encoding 5-lipoxygenase and leukotriene A 4 hydrolase are linked to increased risk of myocardial infarction (Helgadottir et al., 2006; Helgadottir et al., 2004). Some studies have also shown that the 5-lipoxygenase pathway is involved in atherosclerosis (Di Gennaro et al., 2010; Funk, 2005; Qiu et al., 2006a; Spanbroek et al., 2003). As well, plasma LTC 4 levels (Takase et al., 1996) and both CysLT 1 R and CysLT 2 R expression (Jiang et al., 2008) have been shown to increase after myocardial infarction. Previous work by our group has shown that endothelial-specific overexpression of human CysLT 2 R in transgenic mice resulted in significantly greater myocardial infarcts after coronary vessel ligation, which was attenuated by the nonselective cysLT receptor antagonist Bay-u9773 [4-(((1R,2E,4E,6Z,9Z)-1-((1S)-4carboxy-1-hydroxybutyl)-2,4,6,9-pentadecatetraen-1-yl)thio)benzoic acid]. The same study also showed that overexpression of CysLT 2 R resulted in enhanced adhesion molecule up-regulation as well as leukocyte infiltration (Jiang et al., 2008). However, because of the absence of a 45 CysLT 2 R-selective antagonist (Moos and Funk, 2008), the precise pathological contributions of CysLT 1 R and CysLT 2 R could not be dissected (Jiang et al., 2008). Recent breakthroughs in CysLT 2 R-related pharmacology have allowed the characterization of both a CysLT 2 R-selective agonist [N-methyl LTC4 (NMLTC 4 )] (Yan et al., 2011) and antagonist [3-(((3-carboxycyclohexyl)amino)carbonyl)-4-(3-(4-(4(cyclohexyloxy)butoxy)phenyl)propoxy)-benzoic acid (HAMI3379)] (Wunder et al., 2010). In this article, we characterize the pharmacological properties of another CysLT 2 R-selective antagonist 3-(((3-carboxycyclohexyl) amino)carbonyl)-4-(3-(4-(4-phenoxybutoxy)phenyl)propoxy)benzoic acid (BayCysLT 2 ) and use it to investigate the specific role of CysLT 2 R in myocardial injury and vascular leakage. We show that BayCysLT 2 fully attenuates the exacerbation of myocardial infarction injury seen in mice overexpressing CysLT 2 R and accomplishes this by reducing leukocyte infiltration. 46 2.3. Materials and Methods 2.3.1. BayCysLT 2 Synthesis All reagents were purchased from commercial sources and used without further purification. Column chromatography was carried out using Silica-P Flash silica gel (60 Å, 40–63 μm, 500 m2/g) from Silicycle (Quebec City, Canada). 1H (and 13C) NMR spectra were recorded at 25°C on a Bruker Avance Spectrometer (Bruker Daltonics, Billerica, MA) at 300 (75) or 400 (100) MHz. The desired compound, 3-(((3-carboxycyclohexyl)amino)carbonyl)-4-(3-(4-(4phenoxybutoxy)phenyl)-propoxy)benzoic acid (BayCysLT 2 ) is a patented antagonist from Bayer Pharmaceuticals (Montville, NJ) (Harter et al., 2009) and was synthesized according to the route in Figure 2-1 from four commercially available starting materials. 3-(4-hydroxyphenyl) propionic acid (Compound 1 – refer to Figure 2-1), 4-hydroxyisophthalic acid (7), and 3aminocyclohexane-1-carboxylic acid (10) were purchased from TCI America (Portland, OR), and 4-phenoxy-1-butyl bromide (3) was from Alfa Aesar (Ward Hill, MA). After 3 h at room temperature, a solution of 1 (1.5 g, 9.02 mmol) in 15 ml of acidified MeOH (5% final volume with HCl) gave its corresponding methyl ester (2). The solvent was evaporated, and the residue was dissolved in EtOAc. The solution was washed with saturated NaHCO3, dried over MgSO 4 , and concentrated in vacuo to give a yellow oil in quantitative yield. Spectroscopic characterization was consistent with that reported in the literature (Lewin et al., 2005). 47 Methyl-3-(4-(4-phenoxybutoxy)phenyl) Propanoate (4): a solution of 2 (1 g, 6.012 mmol) and 3 (1.5 g, 6.55 mmol) in 15 ml of acetonitrile was mixed with K 2 CO 3 (2.5 g, 2 mmol) and heated to reflux for 15 h. Solvent was removed, and the residue was taken up in EtOAc/H 2 O for workup. The aqueous phase was washed with EtOAc and combined. The organic extracts were washed with brine, dried over MgSO 4 , filtered through a silica plug, and concentrated in vacuo. The residue was purified with flash chromatography using cyclohexane/EtOAc (19:1) to give 1.5 g of 4 (81% yield). Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). 3-(4-(phenoxybutoxy)phenyl)-1-propanol (5): LiAlH 4 (1 M in 5 ml of THF) was introduced dropwise, while stirring, to a solution of 4 (1.3 g, 4.16 mmol) in minimal (~5 ml) THF. The solution was stirred at room temperature for 30 min. MeOH (1–2 ml) was cautiously added to the suspension to destroy excess LiAlH4. The mixture was then added to 1 M HCl and extracted with EtOAc. The organic phase was washed with brine, dried over Na 2 SO 4 , and concentrated to give 5 in quantitative yield. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). 1-(3-bromopropyl)-4-(4-phenoxybutoxy)benzene (6): alcohol 5 (1.4 g, 4.86 mmol) was dissolved in 10 ml of THF, to which was added triphenylphosphine (1 g, 4 mmol) and tetrabromomethane (1.2 g, 4 mmol) with stirring. The reaction became cloudy after 5 min and was complete after 4 h as judged by TLC. The precipitate was filtered off, the filtrate was concentrated in vacuo, and the residue was subjected to flash chromatography on silica gel with 48 cyclohexane/EtOAc (19:1) to yield 1.23 g (3.39 mmol) of 6 in 69% yield. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). 2-hydroxy-5-(methoxycarbonyl)benzoic Acid (8): 4-hydroxyisophthalic acid (7) (1 g, 5.5 mmol) was refluxed in MeOH (10 ml) containing 5% concentrated sulfuric acid (by volume) for 6 h. The mixture was carefully poured into ice water. NaHCO 3 was added to adjust the pH to 8, and the mixture was filtered retaining both the precipitate and filtrate. The precipitate was dissolved in EtOAc and purified by silica gel flash chromatography with 19:1 cyclohexane/EtOAc and trace amounts of glacial acetic acid to separate the desired product from the diester dimethyl 4-hydroxyisophthalate (9), which was the major product at 0.6 g (2.75 mmol). The undesired diester 9 could be converted to the desired monoester by hydrolysis in wet pyridine (10 ml) at reflux for 18 h. The reaction progress was monitored by TLC and purified by silica gel flash chromatography (19:1 cyclohexane/EtOAc) after acidification with HCl, extraction with chloroform, washing with water and NaHCO3, and concentration under reduced pressure. The combined yield of monoester was 80%. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). Methyl 3-aminocyclohexane-carboxylate Hydrochloride (11): 3-aminocyclohexane-1carboxylic acid hydrochloride (10) (1 g, 7.0 mmol) was stirred together with trimethylsilyl chloride (2 ml, 0.012 mmol) in 15 ml of MeOH at room temperature overnight to give a viscous oil, which under vacuum became 0.4 g of a spongy foam in 36% yield. The product was used immediately in the next synthetic step. 49 Methyl 4-hydroxy-3-((3-(methoxycarbonyl)cyclohexyl)amino)carbonyl Benzoate (12): the entire yield of 11, 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (1 g, 6 mmol) and hydroxybenzotriazole (0.5 g, 4 mmol) was dissolved in 15 ml of Et 3 N; this mixture was added dropwise to a solution of 8 in 10 ml of CH 2 Cl 2 . After stirring for 15 h at room temperature, 25 ml of H 2 O was added to the reaction mixture, and the organic phase was extracted. Combined extracts were washed with brine, dried over MgSO 4 , and concentrated in vacuo. The crude product was then purified by flash chromatography on silica gel with cyclohexane/EtOAc (4:1) to yield the product in 63% yield. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). Methyl 3-(((3-(methoxycarbonyl)cyclohexyl)amino)carbonyl)-4-(3-(4-(4-phenoxybutoxy) phenyl)propoxy) Benzoate (13): the coupling product 12 (0.5 g, 1.50 mmol) was dissolved in butylnitrile (25 ml), to which K2CO3 (5 g, 36 mmol) was added and the mixture was heated to reflux. The alkyl bromide 6 (0.6 g, 1.7 mmol) was added over 15 h until 12 was consumed as judged by TLC. A 5% solution of NaH 2 PO 4 (10 ml) was then added to the reaction mixture, and the organics were extracted with EtOAc, washed with brine, dried with MgSO4, and purified by flash chromatography on silica gel with cyclohexane/EtOAc (2:1). Purification yielded 17%. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). BayCysLT 2 : methyl ester 13 (0.15 g, 2.43 mmol) was dissolved in 15 ml of THF and 8 ml of a 1:1 solution of MeOH and 2 M NaOH. The mixture was heated to 60°C for 4 h and then acidified with concentrated HCl, extracted with EtOAc, and precipitated with diethyl ether to 50 obtain the product in quantitative yield. Spectroscopic characterization was consistent with that reported in the literature (Harter et al., 2009). Stock solution (100 mg/ml) was prepared in DMSO for use in all experiments. 2.3.2. β-Galactosidase-β-Arrestin Assay The β-galactosidase-β-arrestin complementation assay for CysLT 2 R activation was described recently (Yan et al., 2011). In brief, Bay-u9773 (Cayman Chemical, Ann Arbor, MI) or BayCysLT 2 (10 pM to 10 μM final concentration) were preincubated for 15 min with C2C12 myoblast cells expressing modified hCysLT 2 R and β-arrestin-2 (25,000 cells/well in 96-well plates; 100 μl) at 37°C. Cells were stimulated with LTD 4 (30 or 300 nM, correlating approximately to the EC 50 and 10× EC 50 values, respectively) (Cayman Chemical) for 1 h after which 50 μl of Tropix Gal-Screen (Applied Biosystems, Foster City, CA) luminescent reagent mix was added to each well. The luminescent readout [relative light units (RLU)] was recorded after incubation at 28°C. In separate assays, C2C12 cells expressing modified hCysLT 2 R and β-arrestin-2 were preincubated with BayCysLT 2 at a fixed concentration (100 nM) for 15 min, then stimulated with varying concentrations of LTD 4 (10 pM to 10 μM) for 1 h. The RLU were recorded after 1h incubation at 28°C. 2.3.3. Measurement of Agonist-Induced Intracellular Calcium Mobilization 51 The coding regions for CysLT 1 R and CysLT 2 R were subcloned into pcDNA3 vectors (Invitrogen, Carlsbad, CA), and HEK293 cells were stably transfected with Lipofectamine 2000 reagent (Invitrogen) following the manufacturer's instructions. Stably transfected cells were selected with G418 (Geneticin) (2 mg/ml; Invitrogen) in Dulbecco's modified Eagle's medium (Millipore Corporation, Billerica, MA) containing 10% fetal bovine serum, 2 mM l-glutamine, and 100 units/ml penicillin/100 μg/ml streptomycin at 37°C in a humidified atmosphere with 5% CO 2 . For confirmation of antagonist specificity, HEK293 cells expressing either CysLT 1 R or CysLT 2 R were plated onto poly-D-lysine-treated, 3-cm, clear-bottom plates at a density of 25,000 cells/plate. Cells were loaded with 3 mM Fura-2-acetoxymethyl ester dye (Invitrogen) and 0.03% Pluronic F-127 solution (Invitrogen) and incubated for 45 min at 37°C. The cells were then washed twice with Hanks' buffered saline solution (Invitrogen) and incubated at 37°C for 15 min in the presence or absence of 300 nM BayCysLT 2 . All cells were then stimulated with 300 nM LTD 4 (Cayman Chemical), and fluorescence at the 340- and 380-nm wavelengths was measured using a Nikon Eclipse TS-100 microscope (Nikon, Melville, NY) and analyzed using Northern Eclipse 7.0 software (EMPIX Imaging Inc., Mississauga, Canada). At the end of the assay period, nonselective calcium release indicating cell viability was confirmed via stimulation with 20 μM cyclopiazonic acid (Sigma-Aldrich, St. Louis, MO). For generation of an antagonist dose-response curve, HEK293 cells were plated into poly-Dlysine-coated 96-well plates at a density of 25,000 cells/well. The next day, cells were incubated for 1 h with Fluo-4 Direct calcium assay reagent (Invitrogen) with 2.5 mM probenecid 52 (Invitrogen). Cells were then incubated in varying concentrations of BayCysLT 2 for 15 min and stimulated with LTD 4 (Cayman Chemical). Maximum fluorescence at 516 nm, indicating the changes in intracellular calcium concentrations, was then measured using a SpectraMax Gemini XS Microplate Spectrofluorometer (Molecular Devices, Sunnyvale, CA). 2.3.4. Animal Models Transgenic human endothelial cell-specific CysLT 2 R overexpressor (EC) mice (Hui et al., 2004) were described previously. The transgenic mice express seven copies of the human CYSLTR2 gene under the control of the Tie2 promoter/enhancer, integrated in a gene-sparse region of chromosome 6. Hemizygous mice were continuously backcrossed with C57BL/6 mice to generate equal numbers of transgenic and littermate wild-type (WT) controls. 2.3.5. Vascular Ear Permeability Assay A vascular ear permeability assay was conducted as described previously (Hui et al., 2004). In brief, mice received either 1) 3 mg/kg BayCysLT 2 diluted in 2.5% DMSO and 97.5% 1× phosphate-buffered saline (PBS) via intraperitoneal injection or 2) no injection 15 min before anesthesia. Mice were anesthetized intraperitoneally with ketamine/xylazine [150 mg/kg ketamine (Vetalar) and 10 mg/kg xylazine (Rompun)] and received 200 μl of 2% Evans blue dye in PBS via intravenous injection through the tail vein once sedated. After 15 min, the right ear was injected intradermally with 5 ng of NMLTC 4 or 5 ng of LTD 4 , both diluted in saline (0.5% EtOH final concentration). The left ear was injected with vehicle (0.5% EtOH in saline). 53 Animals were euthanized 15 min later. A 6-mm biopsy was removed from both ears and soaked in formamide (750 μl) overnight (~18 h) at 55°C, and absorbance of the extracted Evans blue dye at 610 nm was measured with a Varian Cary 50 spectrophotometer (Agilent Technologies, Santa Clara, CA). Readouts were averaged for each experimental group, and the relative change was calculated by comparing ears injected with NMLTC 4 or LTD 4 with vehicle-injected ears. 2.3.6. Ischemia/Reperfusion Surgical Model Ischemia was induced via ligation of the left anterior descending coronary artery (LAD). In brief, mice 14 to 18 weeks of age were administered 20 mg/kg tramadol (Ultram) at least 1 h before surgery. Mice were then anesthetized with 5% isoflurane, intubated, and constantly ventilated (150 breaths/min) with 1 to 5% isoflurane throughout the procedure. An incision was made at the fourth intercostal space, with 50 μl of 50% lidocaine/50% bupivicaine injected subcutaneously along the incision line as an analgesic. The intercostal muscles were cut, and the pericardium and heart were exposed. The pericardium was pulled apart, and 6-0 silk suture (Ethicon, Somerville, NJ) was passed underneath the LAD and surrounding myocardium. Thirty minutes of ischemia was then induced by tightening the suture against a piece of PE-10 tubing placed on top of the LAD and confirmed by visible paling of the affected myocardium. After ischemia, the ligature was loosened to allow reperfusion. The ribs, muscle, and skin layers were closed, isoflurane delivery was stopped, and animals were extubated as soon as they exhibited signs of consciousness. Extubated animals were given 0.5 to 1.0 ml of warm lactated Ringer's solution (Baxter, Mississauga, ON) and returned to their cages once fully mobile. The entire procedure was performed on a heated pad. 54 Mice were divided into three groups: no injection, vehicle injection, and drug injection. BayCysLT 2 (3 mg/kg) in DMSO was diluted in 1× PBS (final concentration 2.5% DMSO) and delivered to the mice via intraperitoneal injection either 45 min before ischemia or 45 min after reperfusion. All surgical procedures and treatment regimens were approved by the University Animal Care Committee at Queen's University and adhered to the guidelines of the Canadian Council of Animal Care. 2.3.7. Analysis of Infarct Volume Mice were euthanized 48 h post-surgery. The heart was fully exposed, the ligature was retightened, and coronary circulation was perfused with 150 to 200 μl of phthalocyanine blue ink dye (Liquitex, Cincinnati, OH) via the aorta to mark the nonrisk area. The heart was then rinsed in ice-cold PBS, blotted dry, wrapped in plastic wrap, frozen for 15 min at −20°C, and cut transversely along the longitudinal axis into six sections using a 1.0-mm Mouse Heart Slicer Matrix (Zivic Instruments, Pittsburgh, PA). These sections were then immersed in 1% 2,3,5triphenyltetrazolium chloride (Sigma-Aldrich) for 15 min at 37°C to demarcate viable and necrotic tissue. Sections were then removed, blotted dry, pressed flat to 1-mm thickness between two pieces of glass, and immersed in water. Sections were then photographed on both sides using a digital camera (Q-Color5; Olympus, Tokyo, Japan). The infarct area (pale white), the area at risk (brick red), the non-risk area (blue), and the total left ventricular area were calculated for both sides of each section using ImageJ software 55 (National Institutes of Health, Bethesda, MD). The areas for each slice were multiplied by the thickness of the slice to obtain a measure of volume. The cumulative volume for all sections for each heart was used for comparisons. As described previously (Jiang et al., 2008), infarct size was calculated as the ratio of the infarct volume to the volume of the risk area. Animals with risk volume in the 35 to 70% range of total left ventricle volume were used as inclusion criteria in the study. 2.3.8. RNA Extraction and Quantitative Real-Time PCR Total RNA from the risk area of the left ventricle was obtained using guanidinium thiocyanatephenol-chloroform extraction (Jiang et al., 2008). In brief, tissue was manually homogenized while immersed in TRI reagent (Sigma-Aldrich). Chloroform (Thermo Fisher Scientific, Ottawa, ON, Canada) was then added, and the suspension was separated into three phases via centrifugation (10 min, 13,000g, 4°C). The clear upper aqueous layer was isolated, and RNA was precipitated using isopropranol, pelleted, and resuspended in diethyl pyrocarbonate-treated double-distilled H 2 O (Invitrogen). RNA quality and quantity were assessed using an Agilent Bioanalyzer (Agilent Technologies) and Nanodrop N-1000 spectrophotometer (Nanodrop, Wilmington, DE), respectively. Total RNA was reverse-transcribed to cDNA using the iScript kit (Bio-Rad Laboratories, Hercules, CA) according to the manufacturer's protocol. Quantitative real-time PCR was performed using a thermal cycler (Applied Biosystems model 7500) with SYBR Green PCR master mix (Bio-Rad Laboratories). GAPDH was used as a control housekeeping gene. Murine Cysltr2 primers were acquired from Invitrogen, and all other primers were acquired from Eurofins MWG Operon (Huntsville, AL). All primer sequences are 56 listed in Table 2-1. Data were calculated using the 2−ΔΔCT method and are presented as foldinduction of transcripts for target genes normalized to Gapdh, with respect to controls. 2.3.9. Myeloperoxidase Assay Neutrophil quantification was indirectly assessed by myeloperoxidase (MPO) activity assay. In brief, mice were subjected to 30 min of ischemia and 48 h of reperfusion as detailed above. Some mice were administered 3 mg/kg BayCysLT 2 45 min post-reperfusion. At endpoint, the ventricles were isolated, flash-frozen, and stored at −80°C. All mice were heparinized before sacrifice. Samples were thawed on ice and placed into 20 mM potassium phosphate (Thermo Fisher Scientific) buffer, pH 7.4, at a 1 mg/20 μl ratio. Samples were then homogenized (PowerGen model 125; Thermo Fisher Scientific) and centrifuged at 10,000g for 5 min at 4°C. The supernatant was discarded, and the pellet was resuspended in 50 mM potassium phosphate (Thermo Fisher Scientific) buffer, pH 6.0, with 0.5% hexadecyltrimethylammounium bromide (Sigma-Aldrich), sonicated (Ultrasonic Dismembrator model 500; Thermo Fisher Scientific), and immersed in a water bath at 60°C for 2 h. Samples were then centrifuged at 10,000g for 5 min at 4°C, and the supernatant was assayed in a 96-well plate by adding 7 μl of resuspended heart tissue to 200 μl of O-dianosidine buffer [0.5 mM O-dianosidine dihydrochloride (SigmaAldrich), 89.95% double distilled H 2 O, 10% 50 mM potassium phosphate buffer, pH 6.0, and 0.05% H 2 O 2 (Sigma-Aldrich)]. The change in absorbance at 460 nm was measured every 30 s for 5 min in a 96-well plate reader (FLUOStar Optima, BMG Tech GmbH, Offenburg, Germany). 57 Gene Gapdh Cysltr2 Vcam1 Icam1 Forward Primer (5′ to 3′) Reverse Primer (5′ to 3′) Amplicon (bp) CATGGCCTTCCGTGTTCCTA TTCCAGAATCAGGTCCACGTG TTGGCTGTGACTCCCCTTCTT ACCACGGAGCCAATTTCTCAT ATGCCTGCTTCACCACCTTCT GCAGTTGCATGGTATGGCAGAA AGAGCTCAACACAAGCGTGGA TGGAAGATCGAAAGTCCGGAG 106 102 104 110 Table 2-1: Primers used in qualitative real-time PCRs. Gene names, primer sequences, and amplicon sizes in base pairs (bp) for primers used in this study are shown. 58 2.3.10. Statistical Analysis Dose-response curve data were analyzed by nonlinear regression using Prism 5.0 software (GraphPad Software Inc., San Diego, CA). Two-tailed unpaired Student's t tests were performed using Prism 5.0 software. All values are given as mean ± SEM. A p value less than 0.05 indicated statistical significance. 59 2.4. Results 2.4.1. Synthesis and chemistry 3-(((3-Carboxycyclohexyl)amino)carbonyl)-4-(3-(4-(4-phenoxybutoxy)phenyl)-propoxy)benzoic acid, termed BayCysLT 2 , was synthesized (as outlined in Figure 2-1) using a similar protocol to that used by Harter et al. (2009). In lieu of purchasing it, we synthesized compound 2 via esterification of 1 as performed by Lewin et al. (2005). Furthermore, in lieu of the literature (Harter et al., 2009), where 8 was purchased, we generated compound 8 by pyridine hydrolysis of the intermediate diester 9 according to Coutts et al. (1981). 1H-NMR, 13C-NMR, and highresolution mass spectra, as well as melting point characterization of our BayCysLT 2 , were consistent with the literature report (Harter et al., 2009). 2.4.2. BayCysLT 2 is more potent than Bay-u9773 A β-galactosidase/β-arrestin assay was used to determine the relative potencies of BayCysLT 2 and the dual CysLT 1 R/CysLT 2 R antagonist Bay-u9773. Cells were preincubated for 15 min before stimulation with either 30 or 300 nM LTD 4 . The IC 50 value for BayCysLT 2 was determined to be 53 nM when cells were stimulated with 30 nM LTD 4 (~EC 50 for LTD 4 in this assay). Increasing the stimulus concentration 10-fold to 300 nM resulted in an increase in the IC 50 to 274 nM. Comparatively, Bay-u9773 was found to have an IC 50 of 4.6 μM when stimulated with 300 nM LTD 4 (Figure 2-2A). Thus, BayCysLT 2 is an approximately 20-fold more potent CysLT 2 R inhibitor, as tested via the β-arrestin assay. 60 Figure 2-1: Chemical synthesis of BayCysLT 2 . Chemical synthesis of BayCysLT 2 . Compounds are indicated by bold numbers and are described under Materials and Methods. Reagents, conditions, and percentage yields are indicated as follows: (a) MeOH, HCl (5%), rt, 3 h, 100%; (b) K 2 CO 3 , MeCN, reflux, 15 h, 81%; (c) LiAlH 4 , THF, rt, 30 min, 100%; (d) Ph 3 P, CBr 4 , THF, rt, 4 h, 69%; (e) MeOH, HCl (5%), reflux, 6 h, 30%: 70%; (f) C 5 H 5 N (aq) , reflux, 18 h, 80%; (g) (CH 3 ) 3 SiCl, MeOH, rt, 18 h, 36%; (h) 1-ethyl3-(3-dimethylaminopropyl) carbodiimide, hydroxybenzotriazole, Et 3 N, CH 2 Cl 2 , rt, 15 h, 63%; (i) K 2 CO 3 , C 5 H 10 N, reflux, 15 h, 17%; (j) NaOH, MeOH, THF, 60°C, 4 h, 100%. 61 We then assessed the competitive nature of BayCysLT 2 inhibition by holding its concentration constant at 100 nM and stimulating the cells with 10 pM to 10 μM LTD 4 . BayCysLT 2 treatment resulted in a rightward shift in the dose-response curve. However, the agonist concentrations used were not potent enough to elicit a plateau response in antagonist-treated cells (preventing the generation of a Schild plot). LTD 4 stimulation-elicited luminescence increased in a dosedependent manner in the presence of BayCysLT 2 (Figure 2-2B). 2.4.3. BayCysLT 2 is selective for CysLT 2 R in vitro and in vivo We sought to confirm BayCysLT 2 selectivity for CysLT 2 R compared with CysLT 1 R. We examined intracellular calcium mobilization in HEK293 cells stably transfected with either CysLT 1 R or CysLT 2 R in response to LTD 4 stimulation and whether BayCysLT 2 could abrogate this response. Both CysLT 1 R- and CysLT 2 R-transfected cells responded robustly to challenge with 300 nM LTD 4 , and although the CysLT 1 R response was transient, the CysLT 2 R response was relatively sustained (Figure 2-3A and C respectively). Application of 100 nM BayCysLT 2 for 15 min before leukotriene stimulation failed to affect intracellular calcium mobilization in CysLT 1 R-transfected cells (Figure 2-3B), but virtually eliminated responses in CysLT 2 Rtransfected cells (Figure 2-3D). Cyclopiazonic acid stimulation at the end of each experiment demonstrated cell viability. Antagonist dose-response curves showed that BayCysLT 2 is ≥500fold more selective for CysLT 2 R than CysLT 1 R in HEK cells stimulated with 300 nM LTD 4 (Fig. 2-4). 62 Figure 2-2: BayCysLT 2 is a more potent antagonist of the human CysLT 2 receptor than Bay-u9773. A, RLU response by C2C12 cells (n = 4) expressing modified hCysLT 2 R and β-arrestin-2 to 30 or 300 nM LTD 4 stimulation in the presence of varying concentrations of BayCysLT 2 or Bayu9773. IC 50 values were determined to be 53 nM (30 nM LTD 4 ) and 274 nM (300 nM LTD 4 ) for BayCysLT 2 and 4.6 μM (300 nM LTD 4 ) for Bay-u9773. B, a constant concentration (100 nM) of BayCysLT 2 was incubated with increasing concentrations of LTD 4 . Response to LTD 4 increased in a dose-dependent manner in the presence of BayCysLT 2 , but did not reach a plateau phase (n = 4). Means of duplicates for each sample are shown (with error bars depicting SEM). 63 Figure 2-3: BayCysLT 2 selectively antagonizes the human CysLT 2 receptor calcium responses. BayCysLT 2 selectively antagonizes the human CysLT 2 receptor calcium responses. Intracellular calcium mobilization in HEK cells expressing either CysLT 1 R (HEK-CysLT 1 R) or CysLT 2 R (HEK-CysLT 2 R) receptor, expressed as the ratio between fluorescence detected at 340 and 380 nm, was elicited via stimulation with 300 nM LTD 4 . Tracings and images depict one cell representative of its experimental group (n = 10–15). Color coding representative of relative intracellular calcium concentrations is indicated on the vertical scale bar, with red being highest and purple the lowest. At the end of each LTD4 stimulation, 20 μM cyclopiazonic acid (CPA) was added to test for nonselective calcium release. A and B, HEK-CysLT 1 R cells responded to leukotriene stimulation both in the absence (A) and presence (B) of 100 nM BayCysLT 2 with a sharp increase in intracellular calcium. C and D, HEK-CysLT 2 R cells responded to leukotriene stimulation in the absence (C) but not in the presence (D) of 100 nM BayCysLT 2 . 64 Figure 2-4: BayCysLT 2 is ≥500-fold more selective for CysLT 2 R than CysLT 1 R. Antagonist dose-response curve for HEK-CysLT 2 R (●) cells was constructed by plotting the fluorescence in response to challenge with 300 nM LTD 4 at each antagonist concentration and used to determine the IC 50 value. HEK-CysLT 1 R (○) cells did not exhibit significant decrease in calcium response to leukotriene stimulation even in the presence of 100 μM BayCysLT 2 . Values are given as mean ± SEM. (n = 3–8). 65 The selectivity of BayCysLT 2 was examined in vivo by examination of vascular leakage in the ear after agonist stimulation. Evans blue dye was injected into the circulation of anesthetized WT (C57Bl/6) and EC transgenic mice, and vascular permeability was assessed by the extravasation of blue dye into the aural tissue in response to cysLT challenge, as measured by absorbance at 610 nm. In WT mice injected with the CysLT 1 R antagonist MK-571 [(E)-3-(((3(2-(7-chloro-2-quinolinyl)ethenyl)phenyl)((3-(dimethylamino)-3-oxopropyl)thio)methyl)thio)propanoic acid], stimulation with LTD 4 resulted in significantly greater dye extravasation than vehicle-injected controls (Figure 2-5A). Mice injected with BayCysLT 2 showed significantly less dye extravasation in both LTD 4 - and NMLTC 4 -treated ears compared with mice treated with MK-571 (Figure 2-5A). In EC mice, LTD 4 injection resulted in significantly greater dye extravasation compared with vehicle control, a phenomenon that was almost eradicated when mice were treated with BayCysLT 2 (Figure 2-5B). 2.4.4. BayCysLT 2 attenuates exacerbation of myocardial damage in EC mice Given our previous findings (Jiang et al., 2008) using the nonselective cysLT receptor antagonist Bay-u9773 in a myocardial ischemia/reperfusion (I/R) injury model, as well as more recent in vitro data with a CysLT 2 R-specific antagonist (Wunder et al., 2010), we sought to determine whether the exacerbation of myocardial damage after ischemia/reperfusion found in EC transgenic mice was CysLT 2 R-dependent. In accordance with previous work, there was no difference in risk volumes between any of the experimental groups (Figure 2-6B). Infarct volume in EC transgenic mice was significantly greater than in WT controls. 66 Figure 2-5: BayCysLT 2 is a potent selective antagonist at the CysLT 2 receptor in vivo. Vascular permeability in murine ears in response to intradermal LTD 4 or NMLTC 4 challenge was measured by quantifying Evans blue dye extravasation and normalizing values against vehicle-injected controls. A, LTD 4 injection caused increased vascular permeability in wild-type mice in the presence of the CysLT 1 R antagonist MK-571, but not in mice treated with BayCysLT 2 . NMLTC 4 injection in BayCysLT 2 -treated mice yielded similar results to LTD 4 injected counterparts. B, LTD 4 injection in EC transgenic mice resulted in increased vascular permeability compared with vehicle-treated controls, and this phenomenon was attenuated by BayCysLT 2 pretreatment. * = p < 0.05. Values are given as mean ± SEM. (n = 4–5). 67 However, treatment with BayCysLT 2 (3 mg/kg) in EC mice 45 min before ischemia attenuated infarct damage. Mice subjected to vehicle injections did not differ in infarction volume compared with sham counterparts. WT mice injected with BayCysLT 2 did not differ in infarction volume compared with vehicle or sham groups (Figure 2-6C). Given that selective CysLT 2 R inhibition in this study and nonselective CysLT 2 R inhibition in a previous study (Jiang et al., 2008) did not affect infarct volume in wild-type mice and myocardial infarction damage occurs in two distinct phases (one consisting of ischemia/reperfusion injury, the second mediated by the inflammatory response to the initial injury), we propose that CysLT 2 R is involved in the second phase. To evaluate this hypothesis, we injected EC transgenic mice with 3 mg/kg BayCysLT 2 45 min after reperfusion. Risk volumes did not differ significantly between experimental groups (Figure 2-6B). However, infarct volume in this group was significantly reduced compared with transgenic mice that did not receive BayCysLT 2 (Figure 2-6C). 2.4.5. Upregulation of adhesion molecule gene expression is attenuated by BayCysLT 2 ICAM-1 and VCAM-1 are essential proteins for leukocyte adhesion and subsequent diapedesis. Indeed, previous work has found both ICAM-1 and VCAM-1 mRNA levels to be significantly elevated in EC mice 3 h after I/R compared with basal levels (Jiang et al., 2008). We report here that ICAM-1 mRNA expression is significantly up-regulated in EC transgenic mice at 48 h postI/R relative to basal levels, and application of BayCysLT 2 45 min 68 Figure 2-6: BayCysLT 2 attenuates CysLT 2 R-mediated exacerbation of myocardial ischemia/reperfusion injury in mice. A, representative 2,3,5-triphenyltetrazolium chloride-stained ventricular sections from WT (white bars) and EC mice (black bars) given no injection (ctrl), vehicle injections (veh), 3 mg/kg BayCysLT 2 45 min before ischemia (drug −45), or 3 mg/kg BayCysLT 2 45 min post-reperfusion (drug +45) are presented. B and C, morphometric analysis of left ventricle volume at risk (B) and infarct size (C) in the groups mentioned above showed that infarction volume was significantly greater in EC mice compared with WT, and administration of BayCysLT 2 either before or after ischemia/reperfusion attenuated this response. * = p < 0.05; **, p < 0.01. Values are given as mean ± SEM. (n = 5–7). 69 Post-reperfusion attenuated this response (Figure 2-7A). Concordantly, VCAM-1 mRNA was also significantly up-regulated in transgenic mice at 48 h post-I/R relative to basal levels. Application of BayCysLT 2 45 min post-reperfusion also attenuated this response (Figure 2-7B). MPO activity was quantified as an indirect assessment of neutrophil presence within the ventricles of mice 48 h post/I/R. MPO activity was significantly increased in EC transgenic mice after I/R compared with wild-type counterparts. However, transgenic mice treated with BayCysLT 2 45 min post-reperfusion showed significantly less MPO activity than untreated transgenic mice (Figure 2-7C). 70 Figure 2-7: BayCysLT 2 attenuates indirect measures of leukocyte recruitment in ischemia/reperfusion injury-induced diapedesis. A and B, EC mice showed significantly increased VCAM-1 (A) and ICAM-1 (B) mRNA expression post-I/R in comparison with non-infarcted controls. Treatment with 3 mg/kg BayCysLT 2 post-I/R resulted in significant attenuation of VCAM mRNA up-regulation. ICAM mRNA up-regulation was also reduced, albeit at borderline significant levels. C, infarcted EC mice also showed significantly increased myeloperoxidase enzyme activity (1 unit is defined as the amount of MPO required to convert 1 μmol of H 2 O 2 in 1 min) present in the myocardial area at risk, indicating an increased presence of neutrophils, compared with infarcted WT animals. Increase in neutrophil presence in the tissue was significantly attenuated by treatment with 3 mg/kg BayCysLT 2 post-I/R. n = 3–4 for mRNA quantification; n = 5 for MPO assay. * = p < 0.05. Values are given as mean ± SEM. 71 2.5. Discussion We hereby demonstrate the in vitro and in vivo characterization of a CysLT 2 R-selective pharmacological antagonist and establish its utility in both cell assays and attenuation of postmyocardial infarction inflammatory response and vascular leakage, respectively. BayCysLT 2 is one of a new family of CysLT 2 R-selective antagonists to be characterized that also includes HAMI3379 (Wunder et al., 2010). BayCysLT 2 differs only in one ring structure from HAMI3379 (benzyl versus cyclohexyl ring). BayCysLT 2 is ~10-fold less potent than HAMI3379 [IC 50 50 to 4.3 nM, respectively, in response to 30 nM LTD 4 (Harter et al., 2009)], but is ~20-fold more potent than the dual antagonist Bay-u9773. In addition, unlike Bay-u9773, BayCysLT 2 does not show partial agonist properties, because intracellular calcium mobilization was not observed during antagonist incubation. Both BayCysLT 2 and HAMI3379 are highly selective for CysLT 2 R, with HAMI3379 showing more than 10,000-fold affinity for CysLT 2 R versus CysLT 1 R (Wunder et al., 2010). Our data indicate that BayCysLT 2 is at least 500-fold more selective for CysLT 2 R and seems to show competitive inhibition, as does HAMI3379 (Wunder et al., 2010). Previous work has shown that the cysLTs mediate vascular permeability in peripheral vasculature and this effect is exacerbated in transgenic CysLT 2 R-overexpressing mice and abolished in CysLT 2 R knockout mice (Hui et al., 2004; Moos et al., 2008). We have confirmed, pharmacologically, that CysLT 2 R mediates this vascular permeability response, because BayCysLT 2 treatment attenuated Evans blue dye extravasation in both WT and EC transgenic animals. The mechanism seems to result from calcium signaling, which in turn regulates 72 transendothelial endocytosis/exocytosis of caveolae-generated vesicles (Moos and Funk, 2008). It was also noted that CysLT 2 R blockage with Bay-u9773 resulted in attenuation of exocytosis but not endocytosis in the vascular endothelium, although this phenomenon may potentially be attributed to the partial agonist properties of Bay-u9773 (Moos et al., 2008). Transgenic overexpression of CysLT 2 R in endothelium has been shown to result in a significant exacerbation of damage after myocardial infarction, as well as increased CD45+ cell infiltration, intermyofibral erythrocyte accumulation, and fluid extravasation (Jiang et al., 2008). Previous work has also noted that basal left ventricular function is unaffected by overexpression of CysLT 2 R in the uninjured myocardium, but post-infarction (2 weeks) remodeling was accelerated in the EC heart (Jiang et al., 2008). In this study, we determined that selective blockade of CysLT 2 R with BayCysLT 2 is capable of abrogating the damage observed in EC transgenic mice. Although the mechanism underlying this effect is not fully clear, our data indicate that it probably involves alterations in the post-reperfusion inflammatory response. Myocardial infarction damage can be loosely separated into three phases: ischemic, reperfusion, and inflammatory (Yellon and Hausenloy, 2007). Ischemic damage occurs during the presence of the coronary circulation blockage and is generally characterized by cellular necrosis. Reperfusion damage occurs at the immediate time when circulation is restored to the myocardium and is characterized by oxidative stress, triggering of pro-apoptotic pathways, and myocardial stunning (Yellon and Hausenloy, 2007). Finally, the damage to the tissue caused by the aforementioned two phases initiates the innate immune response, characterized by increased 73 expression of cell-adhesion molecules, as well as leukocyte recruitment and migration (Yellon and Hausenloy, 2007). Our previous studies observed no reduction in cardiac damage after infarction in mice lacking CysLT 2 R or mice lacking 5-lipoxygenase, the enzyme responsible for leukotriene synthesis, compared with wild-type controls (Jiang et al., 2008). In the current study, BayCysLT 2 treatment attenuated increased infarction damage when applied either before ischemia or after reperfusion. Finally, treatment with BayCysLT 2 prevented the increases in cell-adhesion molecule gene expression and leukocyte infiltration (as measured by MPO assay) into the myocardium that is characteristic of the inflammatory response after acute myocardial infarction. These findings indicate that the cysLTs and their receptors do not play a role in the initial ischemia and/or reperfusion components of the acute myocardial infarction injury response, but rather mediate post-I/R inflammation. Given this evidence, we propose a pathological mechanism where CysLT 2 R activation results in heightened facilitation of diapedesis, which in turn enhances the magnitude of the inflammatory response and results in additional damage to the site of injury. CysLTs required for CysLT 2 R activation can be produced via a transcellular mechanism (Zarini et al., 2009). In the myocardium, cysLT production also probably occurs transcellularly with neutrophils (the first inflammatory cells arriving after reperfusion) being the main LTA 4 source and other cell types (e.g., endothelial cells) converting neutrophil-donated LTA 4 into cysLTs. As such, ischemia/reperfusion tissue damage may result in a positive feedback loop resulting in uncontrolled inflammation. Certainly, endothelial or heterogeneous CysLT 2 R activation has 74 been shown to result in the activation of transcription factors and chemokine genes (Thompson et al., 2008; Uzonyi et al., 2006). Prevention of CysLT 2 R activation via pharmacological antagonism would thus be protective by preventing this positive feedback. Although we have proposed a potential mechanism for CysLT 2 R-mediated exacerbation of the inflammatory response here and from our past studies (Jiang et al., 2008), many questions remain to be investigated and there are limitations to our findings. Precisely how does activation of CysLT 2 R in endothelial cells facilitate this up-regulation of pro-leukocyte adhesion genes? What is the exact interplay between endothelium and infiltrating leukocytes to cysLT generation and activation of the receptor? It is possible that other reactive oxygen species are altered under stress conditions upon CysLT 2 R activation. How does the overexpression of CysLT 2 R in the transgenic model used here translate to human myocardial infarction injury? Future endeavors should be made to determine the long-term effects of CysLT 2 R blockade on cardiac function and remodeling postinfarction in our model and to move beyond mice to humans. In summary, we have characterized, both in vitro and in vivo, a CysLT 2 R-selective pharmacological antagonist, BayCysLT 2 . Along with NMLTC 4 , a novel CysLT 2 R agonist (Yan et al., 2011) and HAMI3379 (Wunder et al., 2010), we now have the required pharmacological tools to fully exploit CysLT 2 R biological signaling in health and disease. 75 2.6. Acknowledgements We thank Matthew Crerar and Christie DeVille for assistance with the calcium imaging experiments; Graham E. Garrett for assistance with the NMR and mass spectrometry analysis; Dr. Yiqun Hui for EC mice; Dr. Alastair V. Ferguson for use of his calcium imaging equipment; and Dr. Michael E. Nesheim for use of his plate reader. This work was supported by the Canadian Institutes of Health Research [Grants MOP-79459, MOP-93689] (to C.D.F.); and a Natural Sciences and Engineering Research Council grant (to D.A.P.). C.D.F. holds a Tier I Canada Research Chair in Molecular, Cellular and Physiological Medicine and is recipient of a Career Investigator award from the Heart and Stroke Foundation of Ontario. D.A.P. holds a Tier II Canada Research Chair in Free Radical Chemistry. 76 Chapter 3: Long-term myocardial remodeling post-ischemia/reperfusion injury is mildly impaired by CysLT 2 R antagonism. 77 3.1. Abstract Cardiovascular disease is a prevalent cause of death today. Cysteinyl leukotrienes (cysLTs) are potent inflammatory mediators involved in cardiovascular disease pathogenesis and severity. In particular, the CysLT receptor 2 (CysLT 2 R) is a mediator of myocardial infarction (MI) injury, as mice with endothelial-targeted CysLT 2 R-overexpression (EC) demonstrate increased damage at 48 hours post-MI. However, the long-term effects of CysLT 2 R activation post-MI are unknown. We examined the effects of CysLT 2 R-overexpression and antagonism on cardiac remodeling in mice following ischemia/reperfusion (I/R) injury. Echocardiography images were used to evaluate dimensional and functional properties of mouse hearts at 2 weeks, 4 weeks, and 12 weeks post-I/R. We observed no major differences in heart dimensions or functional properties between wild-type (WT) or EC mice. However, WT and EC mice treated with the CysLT 2 R-selective antagonist BayCysLT 2 displayed mild ventricular dilatation, increased ventricular mass (WT: 101.9 ± 5.6 mg treated vs. 98.8 ± 4.4 mg untreated; EC: 125.9 ± 6.4 mg treated vs. 106.5 ± 3.7 mg untreated), diminished ejection fraction (WT: 47.8 ± 2.0% treated vs. 56.2 ± 5.1% untreated; EC: 46.1 ± 1.7% treated vs. 55.0 ± 1.7% untreated), and decreased fractional shortening (WT: 23.9 ± 1.1% treated vs. 29.6 ± 3.4% untreated; EC: 22.8 ± 1.0% treated vs. 28.2 ± 1.1% untreated), versus untreated counterparts. Stroke volume, cardiac output, and heart rate were unaffected. These findings indicate that CysLT 2 R antagonism is mildly deleterious to long-term cardiac function post-infarction, despite apparently attenuating acute myocardial injury. This may indicate a necessity for CysLT 2 R signaling for proper wound healing. Further investigation is required to fully understand the role of CysLT 2 R signaling in myocardial injury and remodeling. 78 3.2. Introduction Cardiovascular disease (CVD) is a leading cause of death among both males and females in the industrialized world, and is becoming an increasing problem in the developing world as well. The majority of CVD cases involve myocardial infarction, a condition characterized by blockage of the coronary circulation, resulting in cessation of oxygen and nutrient delivery to the myocardium, leading to tissue death (Yellon and Hausenloy, 2007). Acute treatment entails rapid reperfusion of the myocardium, but that induces a series of responses that result in additional injury to the myocardium - a phenomenon termed "reperfusion injury." Reperfusion injury, in turn, triggers a series of pro-inflammatory responses that facilitate debris removal and proper wound healing if properly regulated (Kempf et al., 2012). However, exacerbation of this inflammatory response results in infarct expansion, resulting in increased myocardial damage (Vanhoutte et al., 2007). In addition, complete attenuation or inactivation of pro-inflammatory mechanisms following ischemic insult leads to deleterious remodeling (Heymans et al., 1999). Thus, understanding and regulating the post-myocardial infarction inflammatory response is critical to maximizing wound healing and preserving cardiac function. Leukotrienes (LTs) are potent pro-inflammatory lipid mediators that are most well-known as mediators of airway smooth muscle constriction (Moos and Funk, 2008). They are synthesized from arachidonic acid and secreted primarily by leukocytes, although leukotriene production can occur in other cell types as well (Folco and Murphy, 2006). The leukotrienes consist of the precursor LTA 4 , the pro-chemotaxtic LTB 4 , and the pro-inflammatory LTC 4 , D 4 , and E 4 termed "cysteinyl leukotrienes" (cysLTs) because of a shared cysteine moiety common to all 79 three molecules (Moos and Funk, 2008). The cysLTs bind to three known receptors, the well characterized cysteinyl leukotriene receptor 1 (CysLT 1 R) and receptor 2 (CysLT 2 R), as well as the recently discovered cysteinyl leukotriene E receptor (GPR99) (Heise et al., 2000; Kanaoka et al., 2013; Lynch et al., 1999). LTC 4 and LTD 4 are agonists for CysLT 1 R and CysLT 2 R, while LTE 4 , the most abundant and stable of the cysLTs, possesses little affinity (Heise et al., 2000; Lynch et al., 1999). However, the situation is reversed for GPR99, which binds LTE 4 with much greater affinity than LTC 4 and LTD 4 (Kanaoka et al., 2013). Research has implicated leukotrienes with a role in cardiovascular disease. Indeed, elevated urinary LTE 4 has been found in individuals post-MI (Carry et al., 1992), and leukotriene synthesis pathway enzymes are expressed in atherosclerotic plaques (Allen et al., 1998). In addition, polymorphisms in the genes encoding 5-lipoxygenase (5-LO), 5-LO activating protein (FLAP), LTA 4 hydrolase (LTA 4 H), and LTC 4 synthase (LTC 4 S) have been linked with increased risk of myocardial infarction and/or stroke (Dwyer et al., 2004; Helgadottir et al., 2006; Helgadottir et al., 2004; Wang et al., 2012). The CysLT 2 R is the predominant isoform expressed in the heart (Heise et al., 2000), and is upregulated following oxygen deprivation in heart, retina, and brain (Barajas-Espinosa et al., 2012; Jiang et al., 2008; Shi et al., 2012a). Mice possessing endothelium-targeted CysLT 2 R-overexpression exhibit damage exacerbation following ischemia/reperfusion, as well as elevated myocardial edema and leukocyte infiltration compared to control mice (Jiang et al., 2008). These effects are attenuated by application of the CysLT 2 R-selective antagonist BayCysLT 2 either prior to ischemia onset or after reperfusion, indicating that CysLT 2 R mediates exacerbation of myocardial infarction damage by regulating the post-ischemia/reperfusion inflammatory response (Ni et al., 2011). 80 Despite our previous work implicating the CysLT 2 R as a mediator of myocardial damage following ischemia, we have very little information on how CysLT 2 R-mediated inflammatory response regulation affects cardiac remodeling and long-term cardiac function. Jiang et al. (2008) demonstrated evidence of accelerated remodeling at 2 weeks post-infarction in the endothelium-targeted CysLT 2 R overexpressing mouse model, but did not investigate longer term effects or whether the phenomenon could be attenuated via CysLT 2 R pharmacological antagonism. As such, we investigate herein the effects of CysLT 2 R overexpression, as well as CysLT 2 R-selective antagonism, on cardiac remodeling and function post-myocardial infarction. 81 3.3. Materials and Methods 3.3.1. Mouse Models All mouse models were generated from a C57/Bl6 genetic background. Transgenic endothelial cell-specific human CysLT 2 R overexpressing mice (EC) were described previously (Hui et al., 2004). These mice express seven copies of the human CYSLTR2 gene under the control of the Tie2 promoter/enhancer, integrated in a gene-sparse region of chromosome 6. 3.3.2. Myocardial Infarction Induction As previously outlined (Jiang et al., 2008), myocardial infarction was induced using in vivo transient surgical ligation of the left anterior descending coronary artery (LAD). Mice were administered 20 mg/kg tramadol (Ultram) at least 1 h before surgery. Mice were then anesthetized with 5% isoflurane, intubated, and constantly ventilated (150 breaths/min) with 1 to 5% isoflurane throughout the procedure. An incision was made at the fourth intercostal space, with 50 μl of 50% lidocaine/50% bupivicaine injected subcutaneously along the incision line as an analgesic. The intercostal muscles were cut, and the pericardium and heart were exposed. The pericardium was pulled apart, and 6-0 silk suture (Ethicon, Somerville, NJ) was passed underneath the LAD and surrounding myocardium. Ischemia was then induced for 30 minutes by tightening the suture against a piece of PE-10 tubing placed on top of the LAD and confirmed by visible paling of the affected myocardium. Following ischemia, the ligature was loosened to facilitate reperfusion. The ribs, muscle, and skin layers were closed, isoflurane delivery was 82 stopped, and animals were extubated as soon as they exhibited signs of consciousness. Extubated animals were given 0.5 to 1.0 ml of warm lactated Ringer's solution (Baxter, Mississauga, ON) via subcutaneous injection and returned to their cages once fully ambulatory. The entire procedure was performed on a heated pad. Selected mice were administered 3 mg/kg of BayCysLT 2 via intraperitoneal injection 45 minutes post-reperfusion. All surgical procedures and treatment regimens were approved by the University Animal Care Committee at Queen's University and adhered to the guidelines of the Canadian Council of Animal Care. 3.3.3. Echocardiography Echocardiography was performed using two-dimensional and Doppler ultrasound using a Vevo 770 in vivo micro-imaging system (VisualSonics, Toronto, ON, Canada). Echocardiograms were obtained from mice at two, four, and twelve weeks post-I/R. In addition, mice treated with BayCysLT 2 were subjected to echocardiography one week prior to induction of I/R. Mice were anaesthetized using 1% isoflurane delivered via nose cone. Hair was removed using a combination of mechanical shaving and chemical depilation, and mice were placed on a heated pad in a supine position. Ventricular dimension measurements were taken using B- and M-mode images obtained at the midpapillary level in the parasternal long- and short-axis views (Figure 31). IVS (interventricular septum), LVID (left ventricular internal dimension), LVPW (left ventricular posterior wall), LVOD (left ventricular outer dimension), apical region wall thickness, and left ventricular volume were measured directly at both systole and diastole (Figure 3-1). Ejection fraction, fractional shortening, left ventricular volume (systole and diastole), 83 Figure 3-1: Echocardiography imaging and left ventricular dimensional measurement methodology. Left ventricular dimensions were measured using images captured via echocardiography. The heart was visualized along both the parasternal long axis and short axis in brightness (B) mode, which was used to obtain measurements of apical ventricular wall thickness. Motion (M) mode images were obtained at the mid-papillary region of the left ventricle, and used to measure the dimensions of the interventricular septum (IVS), left ventricular internal dimension (LVID), and left ventricular posterior wall (LVPW). These measurements were then used in formulae in order to calculate functional parameters of cardiac function. 84 stroke volume, cardiac output, and left ventricular mass were calculated from these measurements (see Appendix I for formulae). Multiple measurements were made for each parameter, with an average of 3-5 cardiac cycles used for all calculations. 3.3.4. Statistical Analysis Data was analyzed by two-way analysis of variance with post-hoc Student's T-test using Prism 5.0 software (GraphPad Software Inc., La Jolla, CA). A p value < 0.05 was determined as statistically significant. All values, unless otherwise mentioned, are presented as mean ± SEM. Due to lack of availability of appropriate pre-I/R echocardiography data for all experimental groups, the 2 week post-surgery timepoint was regarded as baseline for the analysis of changes over time. 85 3.4. Results 3.4.1. Age-related weight gain post-ischemia/reperfusion is unaltered by overexpression of endothelial CysLT 2 R or by BayCysLT 2 treatment. We monitored body weight as a general indicator of health in mice following ischemia/reperfusion injury. At 2 weeks post-I/R, all mice had recovered weight lost due surgical intervention. By 4 weeks, WT mice not treated with BayCysLT 2 (p < 0.05), as well as EC mice treated with BayCysLT 2 (p < 0.01) were significantly heavier than their body weights at the time of surgery. By 12 weeks, all groups were significantly heavier compared to their initial weight prior to LAD ligation (p < 0.01). No differences in body weight were noted between experimental groups at any of the timepoints (Table 3-1). 3.4.2. Post-ischemia/reperfusion pathological cardiac hypertrophy is exacerbated in animals treated with BayCysLT 2 . Myocardial injury, if sufficiently deleterious, results in pathological cardiac hypertrophy and ventricular dilatation either in an attempt to compensate for an inability to respond adequately to increased pressure and/or volume load (Frey et al., 2004). Pathological cardiac hypertrophy can manifest as thickening of the interventricular septum (IVS) and left ventricular posterior wall (LVPW) (pressure overload -mediated) or as thinning of the IVS and LVPW with the enlargement of the left ventricular internal dimension (LVID) (volume overload-mediated). In both cases, pathological cardiac remodeling leads to gradual decrease and loss of cardiac 86 Pre-I/R 2 Weeks 4 Weeks 12 Weeks WT 25.0 ± 1.3 25.1 ± 1.2 26.2 ± 1.1# 33.5 ± 1.2## EC 25.1 ± 1.1 25.3 ± 1.5 25.8 ± 1.6 30.7 ± 2.1## WT-BayCysLT 2 24.9 ± 1.4 25.2 ± 1.5 26.1 ± 1.6 32.6 ± 2.2## EC-BayCysLT 2 26.6 ± 1.4 26.7 ± 1.5 28.8 ± 1.9## 36.1 ± 3.0## Table 3-1: Body weights of mice prior to and following ischemia/reperfusion injury at time of echocardiography. Mouse body weight (in grams) was measured prior to LAD ligation, and again prior to each echocardiography session. Mice in all groups gained significant amounts of weight with age, but no significant differences were noted between groups. # = p < 0.05, ## = p < 0.01 versus pre-I/R timepoint. Values are mean ± SEM, n = 6-10. 87 function (McMullen and Jennings, 2007). We noted no changes in IVS thickness over the time course of the experiment, nor any differences between experimental groups. Likewise, we noticed minimal change in LVPW thickness over time or between experimental groups (Table 32, Figure 3-2). However, LVID at diastole was significantly increased post-I/R over time in EC mice treated with BayCysLT 2 . LVID at systole was significantly increased post-I/R over time in EC, EC-BayCysLT 2 , and WT-BayCysLT 2 groups. In addition to this, EC-BayCysLT 2 mice showed significantly increased LVID during diastole versus untreated EC counterparts at 4 and 12 weeks post-I/R, and both WT-BayCysLT 2 and EC-BayCysLT 2 mice showed significantly increased LVID during systole versus untreated counterparts at 4 and 12 weeks post-I/R. LVOD significantly increased over time during both diastole and systole in EC-BayCysLT 2 mice (but no other groups), and was significantly larger in the EC-BayCysLT 2 group than WT-BayCysLT 2 mice during both diastole and systole as well (Table 3-3) (Figure 3-2). Finally, apical wall thickness was significantly decreased over time during diastole and systole in untreated mice (WT and EC), unchanged in the WT-BayCysLT 2 group, and significantly increased in the ECBayCysLT 2 group. Apical wall thickness at 4 and 12 weeks was also significantly greater in mice treated with BayCysLT 2 versus untreated counterparts. However, no difference was observed between genotypes (Table 3-4). Combined, these results indicate that a mild ventricular dilatation is present in mice treated with BayCysLT 2 (but not untreated mice) - one that is slightly more severe in EC mice than WT mice. However, BayCysLT 2 treatment appears to maintain the acute cardioprotective properties previously detailed (Ni et al., 2011), given that apical region (the region with the highest proportion of area at risk during LAD ligation) wall thickness was significantly increased relative to mice that did not receive BayCysLT 2 . 88 Figure 3-2: Representative images of left ventricular remodeling post-myocardial infarction. Representative M-mode images obtained via echocardiography from untreated WT and EC mice (leftmost two columns) and BayCysLT 2 -treated mice (rightmost two columns) at 2 weeks, 4 weeks, and 12 weeks post-myocardial infarction are presented here, with the left ventricular internal dimension highlighted by white arrows. 89 Pre-I/R 2 Weeks 4 Weeks 12 Weeks IVS diastole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 0.70 ± 0.03 0.78 ± 0.04 0.76 ± 0.06 0.76 ± 0.02 0.76 ± 0.03 0.74 ± 0.02 0.73 ± 0.02 0.80 ± 0.03 0.73 ± 0.02 0.78 ± 0.01 0.75 ± 0.03 0.77 ± 0.03 0.71 ± 0.02 0.81 ± 0.03 IVS systole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 0.96 ± 0.03 1.02 ± 0.04 1.01 ± 0.06 1.00 ± 0.02 0.97 ± 0.05 0.98 ± 0.02 0.98 ± 0.03 1.04 ± 0.03 0.95 ± 0.03 1.01 ± 0.02 1.01 ± 0.05 1.00 ± 0.04 0.88 ± 0.04 1.03 ± 0.04 LVPW diastole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 0.78 ± 0.04 0.82 ± 0.08 0.71 ± 0.03 0.82 ± 0.04† 0.77 ± 0.03 0.82 ± 0.03 0.72 ± 0.02 0.74 ± 0.03 0.68 ± 0.02 0.75 ± 0.02 0.77 ± 0.04 0.76 ± 0.02 0.73 ± 0.03 0.79 ± 0.02 - 1.08 ± 0.07 1.14 ± 0.06 1.08 ± 0.03 1.11 ± 0.03 1.09 ± 0.05 1.08 ± 0.03 1.03 ± 0.04 1.16 ± 0.07 1.00 ± 0.03 1.10 ± 0.04 0.91 ± 0.03* 1.03 ± 0.03 0.99 ± 0.04 1.02 ± 0.04 LVPW systole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 Table 3-2: Left ventricular wall measurements of mice post-I/R injury. Interventricular septum (IVS) and left ventricular posterior wall (LVPW) dimensions at diastole and systole were determined from echocardiography at 2, 4, and 12 weeks following ischemia/reperfusion injury. * = p < 0.05 versus non-drug treated counterparts of the same genotype. † = p < 0.05 versus wild-type counterparts receiving the same drug injection regimen. Values are mean ± SEM, n = 6-10. 90 Pre-I/R 2 Weeks 4 Weeks 12 Weeks LVID diastole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 3.73 ± 0.08 3.70 ± 0.08 3.77 ± 0.04 3.85 ± 0.05 3.89 ± 0.08 3.73 ± 0.05 3.67 ± 0.04 3.80 ± 0.06 3.92 ± 0.13 4.08 ± 0.07*### 3.74 ± 0.04 3.90 ± 0.07 4.00 ± 0.11 4.16 ± 0.20*### LVID systole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 2.70 ± 0.11 2.56 ± 0.14 2.56 ± 0.11 2.59 ± 0.07 2.79 ± 0.12 2.61 ± 0.05 2.51 ± 0.07 2.65 ± 0.15 2.54 ± 0.04 2.81 ± 0.07## 2.94 ± 0.12** 3.04 ± 0.08*# 3.06 ± 0.08***### 3.21 ± 0.17**### - 5.30 ± 0.04 5.27 ± 0.05 5.35 ± 0.07 EC WT-BayCysLT 2 EC-BayCysLT 2 5.17 ± 0.08 5.37 ± 0.11 5.47 ± 0.08 5.31 ± 0.07 5.34 ± 0.07 5.47 ± 0.15 5.30 ± 0.11 5.58 ± 0.07## 5.50 ± 0.05 5.40 ± 0.12 5.73 ± 0.09†## LVOD systole (mm) WT EC WT-BayCysLT 2 - 4.64 ± 0.06 4.81 ± 0.08 4.67 ± 0.06 4.67 ± 0.13 4.83 ± 0.04 4.92 ± 0.06 4.66 ± 0.08 4.74 ± 0.12 4.70 ± 0.08 4.77 ± 0.05 4.77 ± 0.08 5.02 ± 0.07**# 4.87 ± 0.10 5.18 ± 0.08†### LVOD diastole (mm) WT EC-BayCysLT 2 Table 3-3: Left ventricular dimensions of mice post-I/R injury. Left ventricular internal dimension (LVID) and outer dimension (LVOD) sizes at diastole and systole were determined from echocardiography at 2, 4, and 12 weeks following ischemia/reperfusion injury. * = p < 0.05, ** = p < 0.01, *** = p < 0.001 versus non-drug treated counterparts of the same genotype. † = p < 0.05 versus wild-type counterparts receiving the same drug injection regimen. # = p < 0.05, ## = p < 0.01, ### = p < 0.001 versus 2 week timepoint. Values are mean ± SEM, n = 6-10 91 0 Weeks 2 Weeks 4 Weeks 12 Weeks Apex diastole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 0.51 ± 0.02 0.52 ± 0.03 0.53 ± 0.02 0.51 ± 0.04 0.49 ± 0.01 0.47 ± 0.01 0.36 ± 0.03## 0.41 ± 0.03 0.49 ± 0.03** 0.49 ± 0.01* 0.41 ± 0.03# 0.36 ± 0.02# 0.56 ± 0.02*** 0.54 ± 0.02***# Apex systole (mm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 0.56 ± 0.02 0.58 ± 0.04 0.59 ± 0.06 0.48 ± 0.05 0.61 ± 0.04 0.53 ± 0.02 0.38 ± 0.02## 0.41 ± 0.05 0.56 ± 0.02*** 0.58 ± 0.02*** 0.45 ± 0.02# 0.34 ± 0.02# 0.62 ± 0.04** 0.61 ± 0.02***# Table 3-4: Apex wall thickness measurements of mice post-I/R injury. Wall thickness in the apical region of the heart at diastole and systole were determined from echocardiography at 2, 4, and 12 weeks following ischemia/reperfusion injury. * = p < 0.05, ** = p < 0.01, *** = p < 0.001 versus non-drug treated counterparts of the same genotype. # = p < 0.05, ## = p < 0.01 versus 2-week timepoint. Values are mean ± SEM, n = 6-10. 92 3.4.3. Mice treated with BayCysLT 2 show mildly decreased cardiac function. We calculated values for cardiac functional parameters using dimensional measurement data obtained from echocardiography images. Ejection fraction (EF) and fractional shortening (FS) were both significantly decreased in the EC-BayCysLT 2 group versus untreated counterparts. In addition, the WT-BayCysLT 2 group also showed lower EF and FS values than untreated counterparts, although this trend was only significant at the 4 week timepoint. Similarly, while no differences in EF or FS between the experimental groups were observed 2 weeks after infarction, EC-BayCysLT 2 mice showed a significant decline in EF and FS by 4 weeks post-I/R versus 2 week timepoint values, and this continued at the 12 week timepoint. WT-BayCysLT 2 mice showed similar (albeit non-significant) declines in EF and FS over time. No differences were observed between groups or over time in stroke volumes, cardiac outputs, or heart rate (Table 3-5). As such, any maladaptation appears to be relatively mild, as decreases in ejection fraction and fractional shortening have not translated to decreased stroke volume or cardiac output. 3.4.4. Left ventricular volume and mass are increased in BayCysLT 2 -treated mice postischemia/reperfusion injury. We also examined left ventricular volume and mass. No differences between groups were identified at the 2 week timepoint, but the EC-BayCysLT 2 group showed significantly larger LV volume during diastole and systole by the 4 week timepoint, and this trend persisted at the 12 week timepoint. Likewise, untreated EC mice and the WT-BayCysLT 2 group also showed 93 Group 0 Weeks 2 Weeks 4 Weeks 12 Weeks Ejection Fraction (%) WT EC WT-BayCysLT 2 EC-BayCysLT 2 54.1 ± 2.3 59.5 ± 4.0 56.3 ± 4.8 61.9 ± 1.7 55.8 ± 2.8 57.7 ± 1.8 60.1 ± 1.6 62.6 ± 0.8 50.8 ± 3.3* 49.7 ± 2.4**# 56.2 ± 5.1 55.0 ± 1.7 47.8 ± 2.0 46.1 ± 1.7*## Fractional Shortening (%) WT EC 27.6 ± 1.5 WT-BayCysLT 2 31.5 ± 2.7 EC-BayCysLT 2 29.6 ± 3.4 33.0 ± 1.2 29.1 ± 1.8 30.2 ± 1.3 31.6 ± 1.1 33.4 ± 0.59 25.7 ± 2.0* 25.2 ± 1.6**# 29.6 ± 3.4 28.2 ± 1.1 23.9 ± 1.1 22.8 ± 1.0## 34.59 ± 2.59 38.22 ± 1.44 36.85 ± 1.53 35.33 ± 1.97 35.7 ± 0.5 36.2 ± 2.3 35.5 ± 3.3 37.0 ± 2.3 34.7 ± 2.4 37.3 ± 1.9 34.9 ± 2.3 36.9 ± 2.1 14.2 ± 1.4 15.0 ± 0.9 15.5 ± 0.3 14.8 ± 1.4 15.6 ± 1.5 15.7 ± 1.3 15.8 ± 0.7 15.6 ± 1.0 14.9 ± 1.3 16.2 ± 1.4 14.6 ± 0.9 15.3 ± 0.8 417 ± 13 391 ± 18 434 ± 15 444 ± 6 437 ± 9 411 ± 18 427 ± 6 439 ± 15 448 ± 13 423 ± 17 416 ± 10 419 ± 14 Stroke Volume (μl) WT EC WT-BayCysLT 2 EC-BayCysLT 2 30.9 ± 1.7 36.0 ± 2.1 Cardiac Output (ml/min) WT EC WT-BayCysLT 2 12.8 ± 1.1 EC-BayCysLT 2 15.7 ± 1.3 Heart Rate (bpm) WT EC WT-BayCysLT 2 EC-BayCysLT 2 414 ± 20 436 ± 17 Table 3-5: Evaluation of functional measurements in mice post-I/R injury. Ejection fraction, fractional shortening, stroke volume, and cardiac output values were measured determined from echocardiography images obtained at 2, 4, and 12 weeks following ischemia/reperfusion injury according to formulae listed in the Materials and Methods. Heart rate measurements were obtained at the time of echocardiography. * = p < 0.05, ** = p < 0.01, *** = p < 0.001 versus non-drug treated counterparts of the same genotype. # = p < 0.05, ## = p < 0.01 versus 2-week timepoint. Values are mean ± SEM, n = 6-10. 94 significantly increased LV volume during systole at the 12 week timepoint. EC mice treated with BayCysLT 2 presented significantly larger LV volumes (during both diastole and systole) than untreated EC counterparts, and the WT-BayCysLT 2 group showed significantly larger LV volume during systole than untreated WT counterparts. Finally, LV volume during systole was significantly higher in the EC-BayCysLT 2 group versus the WT-BayCysLT 2 group. Left ventricular mass was significantly elevated in the EC-BayCysLT 2 group at the 4 and 12 week timepoints, but none of the other groups showed any significant change over time. The ECBayCysLT 2 group also showed significantly higher LV mass compared to untreated EC counterparts or the WT-BayCysLT 2 group at 4 and 12 weeks post-I/R (Table 3-6). 95 Group 0 Weeks 2 Weeks 4 Weeks 12 Weeks LV volume diastole (μl) WT EC WT-BayCysLT 2 EC-BayCysLT 2 59.1 ± 3.2 59.2 ± 2.8 59.9 ± 1.6 63.6 ± 2.1 66.0 ± 3.1 60.0 ± 2.3 57.7 ± 1.1 61.6 ± 2.4 67.6 ± 4.8 73.6 ± 2.9**### 60.4 ± 1.6 66.6 ± 2.7 70.9 ± 4.3 77.3 ± 2.7*### 27.4 ± 2.7 24.5 ± 3.3 26.2 ± 2.7 24.4 ± 1.7 30.0 ± 2.9 25.4 ± 1.2 23.3 ± 1.2 23.2 ± 1.0 33.5 ± 3.3* 37.5 ± 1.9***†### 26.0 ± 3.6 30.1 ± 1.7## 36.5 ± 2.5*# 41.7 ± 1.7***†### 94.3 ± 5.4 105.0 ± 8.6 96.0 ± 4.9 106.7 ± 4.5 104.2 ± 4.3 101.5 ± 4.0 88.0 ± 2.3 101.2 ± 2.5 96.8 ± 5.1 113.7 ± 3.3**†### 98.8 ± 4.4 106.5 ± 3.7 101.9 ± 5.6 125.9 ± 6.4**††### LV volume systole (μl) WT EC WT-BayCysLT 2 EC-BayCysLT 2 LV mass (mg) WT EC WT-BayCysLT 2 EC-BayCysLT 2 Table 3-6: Left ventricular volume and mass measurements in mice post-I/R injury. LV volume at diastole and systole, as well as LV mass, was determined from echocardiography at 2, 4, and 12 weeks following ischemia/reperfusion injury. * = p < 0.05, ** = p < 0.01, *** = p < 0.001 versus non-drug treated counterparts of the same genotype. † = p < 0.05, †† = p < 0.01 versus wild-type counterparts receiving the same drug injection regimen. ### = p < 0.001 versus 2-week timepoint. Values are mean ± SEM, n = 6-10. 96 2 rm cardiac function via pathological remodeling, thus increasing susceptibility for other conditions such as congestive heart failure (Roger et al., 2012). We have previously outlined a role for CysLT 2 R activation as a mediator of myocardial infarction damage (Ni et al., 2011), and herein present results from the first investigation of long-term effects on the heart following myocardial infarction in CysLT 2 R-altered murine models. Contrary to expectations, we noted very few differences in cardiac size and functional properties between wild-type mice and mice with endothelial-targeted CysLT 2 R overexpression. EC mice had slightly larger (but non-significant) LVID and LVOD sizes versus WT mice, but no differences were noted in IVS or LVPW size, or the functional parameters EF, FS, CO, and SV. This contrasts with previous work by Jiang et al. (2008), where non-significant increases in LVID and significant septal thinning and posterior wall thickening in EC mice were noted. However, this study did not demonstrate significant differences between WT and EC mice in terms of cardiac dimension alterations pre-I/R to post-I/R at two weeks post-myocardial infarction. In addition, the anaesthetic, as well as the method of delivery used during the surgical intervention here, differs from that used by Jiang et al. (2008), which may lead to minor alterations that might have affected our results. Indeed, there is some evidence that tramadol and isoflurane may have mild cardioprotective effects during acute myocardial infarction (Belhomme et al., 1999; Bilir et al., 2007; Zhang and Guo, 2009), and we previously noted that average acute infarct size in EC 97 mice, when using our current surgical protocol, was 47% of area at risk (Ni et al., 2011) whereas Jiang et al. (2008), presented an average infarct size of 56%. This difference may account for the discrepancies in cardiac remodeling noted between our investigation and previous findings. Also, Jiang et al. (2008) only investigated remodeling for a 2 week timeframe post-I/R, so the possibility exists that the differences between WT and EC mice observed in that study may not persist beyond that timeframe. Interestingly, however, we noted that mice treated with the CysLT 2 R-selective antagonist BayCysLT 2 actually presented indicators of maladaptive remodeling following ischemia/reperfusion injury. WT mice treated with BayCysLT 2 show significantly increased LVID and LV volume during systole, and display a trend towards decreased ejection fraction and fractional shortening. The phenomenon is more pronounced in EC mice treated with BayCysLT 2 , which showed increased LVID, LVOD, and LV size during both diastole and systole, along with decreased ejection fraction and fractional shortening post-I/R. Paradoxically, BayCysLT 2 appears to have maintained its ability to reduce infarction size, as the ventricular wall thickness in the apical region (which is most severely affected by LAD ligation-induced ischemia) is significantly greater in mice treated with BayCysLT 2 than untreated counterparts. 3.5.2. Why does CysLT 2 R overexpression not result in a worse long-term outcome? The first question that requires addressing is why do EC mice not present more exacerbated signs of cardiac dysfunction and adverse remodeling post-I/R, given the exacerbation of myocardial damage previously observed in this genotype (Jiang et al., 2008; Ni et al., 2011)? A possible 98 answer may be that cardiac damage suffered during 30 minutes of transient ischemia, as delivered in our model, may not be sufficiently deleterious to evoke the full extent of the remodeling response, and this is masking any differences in damage susceptibility between WT and EC mice. Certainly, the absence of septal or posterior wall thinning (two hallmarks of pathological cardiac dilatation (McMullen and Jennings, 2007)) indicates that the adverse remodeling response is mild at best, and the lack of significant changes in cardiac output and stroke volume supports that as well. Indeed, studies employing permanent LAD ligation to induce ischemia report ventricular wall fibrosis far more severe than anything observed in our studies using transient ischemia (Song et al., 2012; Takemura et al., 2009). Little is known about the resolution pathways for CysLT 2 R-mediated inflammation, and there is some evidence that increased leukocyte influx into the site of injury is not always deleterious. In particular, aged animals and humans both present significantly elevated levels of neutrophil and macrophage infiltration post-injury. However, wound healing in these subjects occurs with significantly less scar tissue formation than in younger counterparts (Eming et al., 2007). Additional investigation is clearly required to further our understanding of how CysLT 2 R affects the post-I/R injury inflammatory response, and how that translates to long-term wound healing and cardiac function. 3.5.3. Why does CysLT 2 R blockade result in an apparently worse long-term outcome? The finding that BayCysLT 2 treated mice presented increased cardiac dilatation appears highly counterintuitive - how and why does the abrogation of infarction damage result in increased pathological hypertrophy? The most likely hypothesis is that CysLT 2 R antagonism, through some unknown mechanism, impedes the wound healing pathways that are required for proper 99 injury resolution. While inhibition of some pro-inflammatory cytokines such as TNFα or IFNγ have been shown to result in accelerated wound healing, inactivation of others, such as MCP-1 and IL-6, result in impairment of the repair mechanism (Eming et al., 2007). In addition, depletion of macrophages or neutrophils resulted in exacerbation of injury and delayed wound repair (Nishio et al., 2008; van Amerongen et al., 2007). Indeed, monocytes and macrophages are integral to proper extracellular matrix remodeling post-infarction (Frantz et al., 2013). The CysLT 2 R expression profile in murine circulatory cells is unknown, but receptor expression has been confirmed on eosinophils, monocytes, macrophages, and platelets (Hasegawa et al., 2010; Heise et al., 2000). However, there has been no investigation as to whether CysLT 2 R is expressed on pro-induction inflammatory or pro-resolution inflammatory cells. This is particularly important with respect to monocytes and macrophages, which display distinct proinflammatory (e.g. Ly-6Chi monocytes (in mice), CD14hi/CD16- monocytes (in humans), and M1 macrophages) and anti-inflammatory phenotypes (e.g. Ly-6Clo monocytes (in mice), CD14lo/CD16hi monocytes (in humans), and M2 macrophages) (Mosser and Edwards, 2008; Nahrendorf et al., 2007). In conclusion, we present here that CysLT 2 R antagonism post-ischemia/reperfusion injury results in mild deleterious pathological remodeling versus untreated counterparts. Further investigation is required to explain why attenuation of myocardial infarction injury results in an adverse remodeling phenotype, but these findings indicate that there may be a role for CysLT 2 R activation in facilitating the healing response as well. 100 3.6. Acknowledgements The authors would like to thank Ms. Kristiina Aase for her expertise and guidance in obtaining echocardiography images. We would also like to thank Drs. Raveen Pal and Amer Johri at Kingston General Hospital for their assistance in interpretation of the echocardiography images. This work was funded by the Canadian Institute for Health Research MOP-68930. C.D.F. holds a Tier I Canada Research Chair in Molecular, Cellular and Physiological Medicine and is recipient of a Career Investigator award from the Heart and Stroke Foundation of Ontario. 101 Chapter 4: Multiple-site activation of the cysteinyl leukotriene receptor 2 is required for exacerbation of ischemia/reperfusion injury 102 4.1. Abstract Transgenic overexpression of the human cysteinyl leukotriene receptor 2 (CysLT 2 R) in murine endothelium exacerbates vascular permeability and ischemia/reperfusion injury. Here, we seek to uncover the underlying mechanisms of CysLT 2 R activation-mediated inflammation and delineate the relative contributions of endogenous murine CysLT 2 R and the transgene-derived receptor. We created a novel mouse (EC/KO) by crossing endothelium-targeted overexpression mice (EC) with CysLT 2 R knockout mice (KO) to isolate the role of endothelial CysLT 2 R in tissue injury. Surprisingly, we discovered that myocardial damage in EC/KO mice was not elevated (24 ± 3% vs. 47 ± 2% EC) following ischemia/reperfusion. To examine potential mechanisms for this finding, we studied vascular permeability and leukocyte recruitment/rolling responses in cremaster vasculature following cysteinyl leukotriene (cysLT) stimulation. Mice possessing transgenic endothelial CysLT 2 R overexpression (EC or EC/KO), when stimulated with cysLTs, exhibited vascular hyperpermeability, decreased leukocyte flux, and transient increase in slow-rolling leukocyte fraction. Mice lacking endogenous CysLT 2 R (both KO (20 ± 3 cells/min) and EC/KO (24 ± 3)) showed lower rolling leukocyte flux than WT (38 ± 6) and EC (35 ± 6) mice under unstimulated conditions. EC/KO mice differed from EC counterparts in that vascular hyperpermeability was not present in the absence of exogenous cysLTs. These results indicate that endothelial and “non-endothelial” CysLT 2 R niches have separate important roles in mediating inflammatory responses. Endothelial receptor activation results in increased vascular permeability and leukocyte slow-rolling, facilitating leukocyte transmigration. Other receptors – likely located on resident/circulating leukocytes – facilitate endothelial receptor activation and 103 modulate leukocyte transit. Activation of both receptor location subsets facilitates injury exacerbation. 104 4.2. Introduction Myocardial infarction (MI) is one of the leading killers in the Western world today, and everincreasing obesity rates in the industrialized world makes cardiovascular disease a growing threat to public health (White and Chew, 2008). MI is characterized by a blockage of the coronary circulation, usually caused by atherosclerotic plaque rupture, leading to cessation of oxygen and nutrient delivery to the myocardium. Timely restoration of flow to the affected area, reperfusion, is paramount. However, this treatment method is also a double-edged sword, while necessary to salvage cardiac function, it also evokes "reperfusion damage," a combination of oxidative stress and inflammation that results in additional myocardial injury (Yellon and Hausenloy, 2007). Leukotrienes are potent pro-inflammatory lipid mediators synthesized from arachidonic acid via the actions of 5-lipoxygenase and 5-lipoxygenase-activating protein. Cysteinyl leukotrienes (cysLTs) are a subfamily of leukotriene molecules, so termed due to a common cysteine moiety in their respective structures. The cysLTs, leukotriene C 4 , D 4 , and E 4 , exert their effects by binding to G-protein coupled receptors: cysteinyl leukotriene receptor 1 (CysLT 1 R), cysteinyl leukotriene receptor 2 (CysLT 2 R), and a novel CysLT E receptor GPR99 (Kanaoka et al., 2013). CysLT 1 R binds LTD 4 preferentially to LTC 4 , whereas CysLT 2 R binds LTC 4 and LTD 4 with equal affinity. Both receptors bind LTE 4 , the most stable and abundant cysLT, with weak affinity (Maekawa et al., 2008). In contrast, CysLT E R displays preferential binding affinity for LTE 4 , with weak affinity for LTC 4 and LTD 4 (Kanaoka et al., 2013). 105 CysLT 1 R has been studied extensively and is involved in airway inflammation. Indeed, CysLT 1 R pharmacological antagonists are popular anti-asthma treatment options. CysLT 2 R, in humans, is expressed in umbilical vein endothelial cells, macrophages (Lotzer et al., 2007; Sjostrom et al., 2003), platelets (Hasegawa et al., 2010), the cardiac Purkinje system (Heise et al., 2000), and coronary endothelial cells (Hui et al., 2001). CysLT 2 R research had been previously hampered by a lack of selective pharmacological agents (Moos and Funk, 2008) – the majority of work instead using the non-selective CysLT antagonist/partial agonist Bay-u9773 (Jiang et al., 2008). However, recent studies have characterized two novel CysLT 2 R-selective antagonists: HAMI3379 (Wunder et al., 2010) and BayCysLT 2 (Carnini et al., 2011; Ni et al., 2011; Qi et al., 2011), that are beginning to prove useful as tools for eicosanoid-related research. There is some evidence linking cysLTs to cardiovascular and inflammatory disease. Expression of components of the leukotriene synthesis pathway is found in human atherosclerotic lesions (Spanbroek et al., 2003). Furthermore, genetic polymorphisms in 5-lipoxygenase pathway genes have been correlated to increased MI and stroke risk in some populations (Helgadottir et al., 2005) but not all (Bisoendial et al., 2012; Maznyczka et al., 2008). CysLT 2 R can mediate vascular permeability (Hui et al., 2004; Moos et al., 2008), susceptibility to gastrointestinal tract inflammation (Barajas-Espinosa et al., 2011), and increased vulnerability to ischemic injury in the heart (Jiang et al., 2008) and brain (Qi et al., 2011; Shi et al., 2012b). CysLT 2 R expression is increased following hypoxic/ischemic stress (Barajas-Espinosa et al., 2012; Fang et al., 2007; Jiang et al., 2008; Shi et al., 2012b). Previous work from our laboratory has shown that CysLT 2 R-selective antagonism can abrogate myocardial ischemia/reperfusion injury in transgenic mice overexpressing the human CysLT 2 R receptor in vascular endothelium (EC). 106 However, we were unable to delineate the relative roles of the transgenic endothelial CysLT 2 R versus endogenous murine CysLT 2 R present on various cell types. In order to study this, we have created a novel mouse model by crossing CysLT 2 R knockout mice with EC mice, resulting in a model with selective endothelial CysLT 2 R expression. Using these novel mice, we examine how CysLT 2 R mediates post-ischemic myocardial injury and vascular inflammatory responses. 107 4.3. Materials and Methods 4.3.1. Mouse Models All mice used in this study were on a C57Bl/6 genetic background. EC transgenic mice were previously described (Hui et al., 2004). Briefly, these mice possess 7 copies of the human CYSLTR2 coding region placed under the control of the Tie2 promoter/enhancer for endothelialtargeted CysLT 2 R overexpression. LacZ-CysLT 2 R knockout (KO) mice (Moos et al., 2008) contain a LacZ cassette under control of the Cysltr2 promoter with Cysltr2 disruption. X-Gal staining can be used as a reporter for native sites of CysLT 2 R expression (Moos et al., 2008). These two strains were crossed to generate a whole-body (including endogenous endothelial expression) CysLT 2 R knockout mouse that possesses endothelium-targeted overexpression of the human CysLT 2 R. This mouse is referred to as EC/KO. Details of the endogenous and transgenic CysLT 2 R expression profile of each genotype utilized can be found in Table 4-1. 4.3.2. Myocardial Ischemia/Reperfusion Myocardial infarction was induced via non-permanent left anterior descending coronary arterial ligation, as previously outlined (Ni et al., 2011). Briefly, analgesia was administered (20 mg/kg tramadol (Ultram; Chiron AS, Trondheim, Norway)) at least 1 h prior to surgery in mice 14-18 weeks of age. Mice were then anesthetized with 5% isoflurane, intubated, and constantly ventilated (150 breaths/min) with 1 to 5% isoflurane throughout the procedure. An incision was made at the fourth intercostal space, with 50 μl of 50% lidocaine/50% bupivicaine injected 108 Genotype Endothelial Expression Non-Endothelial Expression WT mCysLT 2 R + mCysLT 2 R + EC mCysLT 2 R + hCysLT 2 R +++ mCysLT 2 R + KO None None EC/KO hCysLT 2 R +++ None Table 4-1: CysLT 2 R expression profile in featured mouse genotypes. Expression levels (denoted by + signs) of endogenous murine and transgenic human CysLT 2 R in endothelial and non-endothelial cells in various murine genotypes. 109 subcutaneously along the incision line as an analgesic. The intercostal muscles were cut in order to expose the heart. The pericardium was pulled apart, and 6-0 silk suture (Ethicon, Somerville, NJ) passed underneath the LAD and surrounding myocardium. Ischemia was induced for 30 minutes by tightening the suture against a piece of PE-10 tubing placed on top of the LAD and confirmed by visible paling of the affected myocardium and/or visibly altered ventricular contraction rhythm. Removal of the tubing and loosening of the ligature allowed reperfusion. The surgical site was closed and mice were extubated as soon as they exhibited signs of consciousness, followed by subcutaneous administration of 0.5 to 1.0 ml of warm lactated Ringer's solution (Baxter, Mississauga, ON), and returned to their cages once fully mobile. The entire procedure was performed on a heated pad. All surgical procedures and treatment regimens were approved by the University Animal Care Committee at Queen's University and adhered to the guidelines of the Canadian Council of Animal Care. 4.3.3. Analysis of Infarct Volume Infarct volume was analyzed 48 hours post surgery as previously described (Ni et al., 2011). The heart was excised and retrogradely perfused via the aorta with phthalocyanine blue ink dye (Liquitex, Cincinnati, OH) following suture re-tightening in order to demarcate the non-risk region. The heart was then rinsed in ice-cold PBS, blotted dry, frozen in plastic wrap at -20°C for 15 minutes, and cut transversely into six 1.0 mm sections. Sections were immersed in 2,3,5triphenyltetrazolium chloride (1%; Sigma-Aldrich) for 15 min at 37°C to demarcate viable and necrotic tissue. Stained sections were photographed on both sides using a digital camera (QColor5; Olympus, Tokyo, Japan). The infarct area (pale white), the area at risk (brick red), the 110 non-risk area (blue), and the total left ventricular area were calculated for both sides of each section using ImageJ software (National Institutes of Health, Bethesda, MD). As described previously (Jiang et al., 2008; Ni et al., 2011), infarct size was calculated as the ratio of the infarct volume to the volume at risk. Animals with risk volume in the 35 to 70% range of total left ventricle volume were used as inclusion criteria in the study. 4.3.4. Intravital Microscopy of the Cremaster Vasculature Vascular permeability and leukocyte recruitment were observed using intravital confocal microscopy. Male mice 12-16 weeks of age were anaesthetized with ketamine (150 mg/kg) and xylazine (10 mg/kg). A catheter was placed in the right jugular vein for experiments that required intravenous fluorescein isothiocyanate (FITC)-labeled albumin injection. The cremaster muscle was then exposed as described previously (McCafferty et al., 2002). Post-capillary venules (20-50 μm) were visualized using a Quorum WaveFX-X1 spinning disk confocal system (Quorum Technologies Inc., Guelph, ON, Canada). Recordings were taken with a Hamamatsu EM-CCD camera (model 09100-13; Hamamatsu Photonics, Hamamatsu, Japan) using MetaMorph software (Molecular Devices, Sunnyvale, CA). In order to assess vascular permeability, FITC-labeled albumin (25 mg/kg body weight) was injected via the jugular catheter, and fluorescence intensity in the preparation was recorded for 1 minute. At this point, cysLTs (5 μmol/L each LTC 4 and LTD 4 ) were administered to the superfused vessel – following which fluorescence readings were recorded for a further 14 minutes. 111 To evaluate leukocyte flux in the cremaster vasculature, mice were prepared as above without catheterization. Bright-field images were recorded for 2 min prior to cysLT administration, following which bright-field segments were recorded at 5, 10, and 15 minutes post-cysLTs for 2 min. No difference in vessel diameter was noted between experimental groups (data not shown). 4.3.5. Analysis of Vascular Permeability Vascular permeability was assessed by measuring FITC-albumin extravasation from the vasculature into the surrounding tissue. Fluorescence intensity, ranging from 0 to 65536 arbitrary units, was measured at five 1000 μm2 sites surrounding the post-capillary venule using MetaMorph software. The absolute fluorescence intensity was recorded every 5 frames, with measurement areas manually adjusted to account for field-of-view drift. The first derivative of the absolute fluorescence intensity was calculated to determine the fluorescence intensity rate of change (a measurement previously termed LIFT – leakage intensity factor for tissues (Moos et al., 2008)). 4.3.6. Analysis of Vascular Leukocytes Bright-field recordings of post-capillary venules (20-50 μm) in the cremaster muscle were taken pre/post-leukotriene stimulation. Two min sequences were recorded prior to leukotriene stimulation and at 5, 10, and 15 min following stimulation. Rolling leukocyte flux, defined as the number of rolling leukocytes/min in the observed vessel (Ley, 1994), was visually analyzed 112 by three observers blinded to the study group using VirtualDub version 1.9.9. Leukocyte velocity of 15-30 randomly selected cells was measured by tracking individual leukocytes/mouse/time point using Image-Pro Plus (Media Cybernetics Inc., Rockville, MD). Median leukocyte velocity was used as a representative indicator of flow speed, as average velocity would be improperly skewed by loosely adherent fast rolling leukocytes. Rolling leukocyte speed distribution was analyzed and presented using histograms (Jung and Ley, 1999). 4.3.7. Complete Blood Count Blood was extracted from mice using a heparinized 25G needle via cardiac puncture immediately following CO 2 -asphyxiation and was stored in 1.5 ml eppendorf tubes. Samples were kept on ice until analysis using a scil Vet abc machine (scil Vet Novations; Barrie, ON, Canada). 4.3.8. Statistical Analysis For each experimental group, the mean and standard error were calculated. To compare groups at a single timepoint, we performed a two-tailed unpaired Student’s t-test. To compare timepoints within a single group, we performed a two-tailed paired Student’s t-test. To compare groups over multiple time-points, we performed one way analysis of variance with post hoc Newman-Keuls t-tests. Comparison of rolling leukocyte velocity between groups was done using a nonparametric Kruskal-Wallis one-way ANOVA to determine significant differences, followed by Dunn's test for multiple comparisons. All statistical analysis was carried out using 113 Prism 5.0 (GraphPad Software Inc., San Diego, CA). p < 0.05 was considered a statistically significant difference. 114 4.4. Results 4.4.1. Endothelium-targeted CysLT 2 R overexpression does not exacerbate infarction damage in the absence of endogenous CysLT 2 R Our previous findings established that transgenic CysLT 2 R overexpression in the endothelium, in the presence of endogenous murine CysLT 2 R expression, results in significantly increased infarction volume following myocardial ischemia/reperfusion in mice (Jiang et al., 2008; Ni et al., 2011). However, transgenic mice without endogenous CysLT 2 R expression (EC/KO) did not show this phenomenon (Figure 4-1). Indeed, both WT (23.2 ± 2.2%, n = 13) and EC/KO (24.0 ± 3.0%, n = 14) groups showed significantly lower infarction volumes than EC mice (47.2 ± 1.9%, n = 12; p < 0.001 against both groups). These findings were unexpected because endogenous CysLT 2 R expression is significantly less than transgene expression (Hui et al., 2004). Volume at risk did not vary between any of the experimental groups (Figure 4-1B). We also found that CysLT 2 R knockout mice (17.8 ± 2.2%, n = 12) trended towards lower infarction volumes than WT and EC/KO groups (Figure 1C; p = 0.096 vs. WT, p = 0.11 vs. EC/KO). These results indicate that exacerbation of CysLT 2 R-mediated myocardial ischemia/reperfusion injury is not solely mediated by transgene-targeted CysLT 2 R. 4.4.2. Vascular permeability responses in EC/KO mice differ from those in EC mice 115 Given that CysLT 2 R-mediated myocardial injury appears not to be exclusively mediated by the endothelium-overexpressed CysLT 2 R transgene, we sought to investigate how and to what extent 116 Figure 4-1: Myocardial injury is not elevated in EC/KO mice following ischemia/reperfusion. (A) Representative tetrazolium chloride-stained ventricular sections from wild-type (WT), Endothelium-targeted CysLT 2 R-overexpressing (EC), CysLT 2 R-knockout (KO), and EC/KO mice. Morphometric analysis of (B) LV volume at risk (C) and infarct size in the groups mentioned above showed that infarction volume was significantly greater in EC mice compared to WT. However, this phenomenon was not seen in EC/KO mice. * = p < 0.05; ** = p < 0.01; values given as mean ± SEM, n = 12-14. 117 knockout of endogenous CysLT 2 R would affect endothelial/vascular permeability. This is especially pertinent as based on previous work (Jiang et al., 2008; Ni et al., 2011) CysLT 2 Rmediated endothelial permeability facilitates tissue injury by permitting leukocyte extravasation, inflammation, and edema. We examined FITC-albumin extravasation from post-capillary venules in the cremaster muscle both before and after exogenous leukotriene stimulation. Minimal extravascular fluorescence accumulation prior to leukotriene stimulation was observed in WT, KO, and EC/KO mice. However, EC mice showed significantly higher absolute fluorescence accumulation (Figure 4-2B) and rate of fluorescence accumulation (Figure 4-2C) as previously determined, which we surmised was due to surgery-induced cysLT release from resident tissue macrophages/mast cells activating high numbers of endothelial cell-expressed CysLT 2 Rs (Moos et al., 2008). Administration of cysLTs resulted in an increase in absolute FITC-albumin extravasation in all experimental groups (Figure 4-3A). EC and EC/KO groups demonstrated increased total leakage and rate of leakage versus the WT group post-cysLTs, while cysLT-challenged KO mice showed significantly less FITC-albumin extravasation compared to WT mice (Figures 4-3B and 3C). Interestingly, EC/KO mice, despite possessing identical transgenic overexpression of CysLT 2 R in the endothelium as EC mice, only showed elevated extravascular FITC-albumin accumulation versus WT mice following cysLT stimulation. This is in contrast to EC mice, which showed increased FITC-albumin extravasation compared to WT mice both prior to and after cysLT stimulation. 118 Figure 4-2: Vascular permeability post-surgical intervention (“basal”) is elevated in EC mice, but not in EC/KO mice. (A) Intravital microscopy recordings of vascular leakage in cremaster post-capillary venules. Representative images of cremaster preparations from mice possessing (from left to right) normal (WT), endothelium-elevated (EC), no (KO), or endothelium-only (EC/KO) CysLT 2 R expression is depicted. Upper panels show fluorescence recordings at the start of the experiment (t = 0); lower panels show fluorescence recordings 1 min later. Scale bar = 50 μm. (B) Absolute change and (C) rate of change in extravascular fluorescence were elevated in EC mice. * = p < 0.05; values given as mean ± SEM, n = 6-7. 119 Figure 4-3: CysLT 2 R activation mediates vascular hyperpermeability. (A) Intravital microscopy recordings of vascular leakage in cremaster post-capillary venules following administration of 5 μmol/L cysLTs. Representative images of cremaster preparations from mice possessing (from left to right) normal (WT), endothelium-elevated (EC), no (KO), or endothelium-only (EC/KO) CysLT 2 R expression is depicted. Upper panels show fluorescence recordings at the start of the experiment (t = 120 seconds); lower panels show fluorescence recordings at the end of the experiment (t = 840 seconds). Scale bar = 50 μm. (B) Representative measurements of absolute fluorescence intensity in cremaster muscle preparations from WT, EC, KO, and EC/KO mice used to calculate FITC-albumin leakage. (C) Rate of extravascular fluorescence accumulation was elevated in all mice following cysLT stimulation, but EC and EC/KO mice showed the greatest increase. * = p < 0.05; ** = p < 0.01; values given as mean ± SEM, n = 5-7. 120 4.4.3. Genetic knockout of endogenous CysLT 2 R reduces basal rolling leukocyte flux in cremaster vasculature. The endothelium also plays a large role in regulating leukocyte capture and transmigration during inflammatory responses (Rao et al., 2007), and we have previously reported increased leukocyte adhesion molecule mRNA expression in EC mice following ischemia/reperfusion injury (Jiang et al., 2008; Ni et al., 2011). In order to examine whether CysLT 2 R expression modulation affects leukocyte recruitment and behaviour, we examined leukocyte flux under basal and cysLT-stimulated conditions in the cremaster vasculature. Rolling leukocyte flux (the number of rolling leukocytes passing through the vasculature/minute) was higher in mice expressing endogenous CysLT 2 R (WT: 38 ± 6 cells/min, n = 12; EC: 35 ± 6, n = 16) than mice lacking endogenous receptor (KO: 20 ± 3, n = 16, p < 0.01 vs. WT, p < 0.05 vs. EC; EC/KO: 24 ± 3, n = 14, p < 0.05 vs. WT) mice under basal conditions (Figure 4-4). Mice that possessed transgenic endothelium-targeted CysLT 2 R overexpression did not show significantly altered rolling leukocyte counts versus non-transgenic counterparts. Complete blood counts for all genotypes were undertaken and no significant differences in baseline leukocyte numbers or differential counts were detected. 4.4.4. Endothelial-targeted CysLT 2 R overexpression results in diminishing leukocyte flux in cremaster vasculature following leukotriene stimulation Next, we investigated the effects of cysLT stimulation on rolling leukocyte flux in murine cremaster post-capillary venules. Following cysLT stimulation, WT mice (n = 6) showed a non- 121 Figure 4-4: Basal rolling leukocyte flux is mediated by endogenous CysLT 2 R. Number of rolling leukocytes/min was counted in WT, EC, KO, and EC/KO mice. WT and EC mice showed significantly elevated rolling leukocyte flux compared to KO and EC/KO counterparts. * = p < 0.05; ** = p < 0.01; values given as mean ± SEM, n = 12-16. 122 significant increase in rolling leukocytes after 5 min (+13 ± 5 cells/min, p = 0.07 vs. start), and this was maintained until the end of the experiment at 15 minutes (+12 ± 5, p = 0.07 vs. start). CysLT-stimulated KO mice cremaster preparations (n = 6) did not show significant changes in rolling leukocytes throughout the duration of the experiment. However, cysLT stimulation of the cremaster vasculature in mice possessing transgenic endothelial CysLT 2 R overexpression resulted in a decrease in rolling leukocytes. Rolling leukocyte count per minute dropped in EC mice (n = 9) by 5 min (-8 ± 5), and had significantly decreased by the 15 minute timepoint (-13 ± 6, p = 0.04 vs. start). Rolling leukocyte flux in EC/KO mice (n = 7) was not changed at 5 min (2 ± 2), but showed a significant decline by 15 min (-10 ± 3, p = 0.002 vs. start). Change in rolling leukocyte flux in EC and EC/KO mice were statistically significant versus WT mice (Figure 4-5). These results indicate that endothelial CysLT 2 R overexpression significantly alters rolling leukocyte flux counts post-leukotriene stimulation. 4.4.5. Cysteinyl leukotriene stimulation results in increased leukocyte slow rolling fraction in cremaster vasculature in mice expressing endothelial CysLT 2 R. Prior to firm adhesion and transmigration, leukocytes undergo “slow rolling,” which is characterized by leukocyte velocity decreasing to less than 10 μm/s. Slow rolling is mediated by E-selectin/CD18 binding, and is triggered by pro-inflammatory cytokines (Jung and Ley, 1999; Jung et al., 1998). We examined rolling leukocyte velocity in cremaster venules in the absence and presence of cysteinyl leukotriene stimulation. Rolling leukocyte velocity distribution in the unstimulated cremaster muscle did not differ significantly between the four genotypes. However, WT (n = 6 mice, 58% 79%), EC (n = 8 mice, 65% 78%), and EC/KO (n = 7 123 Figure 4-5: Rolling leukocyte flux diminishes in mice expressing elevated endothelial CysLT 2 R post-leukotriene stimulation. (A) Representative images of rolling leukocyte flux in cremaster post-capillary venules of WT, EC, KO, and EC/KO mice prior to (left) and 15 min after 5 μmol/L cysLT stimulation (right). (B) Average change in rolling leukocyte flux in mice post-stimulation with 5 μmol/L cysLTs. WT mice show increased numbers of rolling leukocytes, whereas EC and EC/KO mice show significant decline in leukocyte flux. * = p < 0.05; ** = p < 0.01; values given as mean ± SEM, n = 6-9. 124 mice, 59% 70%) groups all presented a significant shift in rolling leukocyte velocity distribution towards increased proportions of slow-rolling cells at 5 min post-cysLT stimulation (Figure 4-6). KO mice (n = 6 mice, 61 64%) did not display this trend. The presence of this phenomenon in EC/KO mice indicates that it is the endothelial CysLT 2 R niche that drives promotion of slow-rolling leukocytes. Interestingly, while WT mice showed a significantly increased proportion of slow-rolling leukocytes through all three post-stimulation timepoints, both EC and EC/KO mice saw slow-rolling cell fraction return to pre-stimulation levels by 10 min (Figure 4-6). Median rolling leukocyte velocity in the unstimulated cremaster muscle did not differ significantly between the four genotypes. However, median rolling leukocyte velocity in WT, EC, and EC/KO mice all decreased significantly at 5 min post-leukotriene stimulation. This phenomenon persisted in WT and EC mice at 10 and 15 min (albeit non-significantly in the latter group), but was transient in EC/KO mice. In addition, median velocity in EC mice was significantly lower than EC/KO mice at 10 min post-cysLT stimulation, and both WT and EC mice showed significantly lower median velocity compared to EC/KO mice at 15 min poststimulation (Figure 4-7). This indicates that activation of CysLT 2 R in other locations apart from the endothelium may also play a role in mediating/sustaining leukocyte slow-rolling. 125 Figure 4-6: Rolling leukocyte velocity distributions in mice before and after cysteinyl leukotriene stimulation. Histograms of rolling leukocyte velocities in murine cremaster vasculature prior to and following application of 5 μmol/L cysLTs. Velocity of individual leukocytes from 6-9 mice (roughly equal numbers of leukocytes were tracked in each mouse) were calculated and the results compiled (n values indicate total number of cells tracked). WT (A-D) mice showed persistent and significant increase in slow-rolling (≤10 μm/s) leukocyte fraction following cysLT application. EC (E-H) and EC/KO (M-P) mice showed a significant increase in slow-rolling leukocyte fraction at 5 minutes that disappeared by the 10 minute mark. KO (I-L) mice did not show any significant change in leukocyte rolling velocity distribution. ** = p < 0.01; *** = p < 0.001 versus untreated. 126 Figure 4-7: Rolling leukocyte velocity medians in mice before and after cysteinyl leukotriene stimulation. Boxplot shows median (heavy line), 25th and 75th percentiles (top and bottom of box respectively), and range of measured velocities (5th to 95th percentile), excluding calculated outliers and extremes (whiskers). Median velocity was significantly decreased in WT (A), EC (B), and EC/KO (D) mice post-leukotriene stimulation. While this phenomenon persisted for the duration of the experiment in WT mice (A), EC mice showed non-significantly decreased median velocity relative to 0 min at 10 and 15 min (B). EC/KO mice showed a return to prestimulation leukocyte median velocity at 10 and 15 min post-stimulation (D). No change in median velocity was noticed in KO mice (C). In addition, significant differences in median velocity were noted between genotypes as well. ** = p < 0.01; *** = p < 0.001 vs. 0 min, # = p < 0.05; ## = p < 0.01 vs. EC group, † = p < 0.05; †† = p < 0.01 vs. WT group. 127 4.5. Discussion 4.5.1. Endothelial CysLT 2 R is not the sole mediator of CysLT 2 R-mediated ischemia/reperfusion injury exacerbation Endothelial expression of functionally-relevant CysLT 2 Rs has been well established for almost a decade now (Hui et al., 2004; Spanbroek et al., 2003), and endothelial-targeted overexpression of CysLT 2 R results in exacerbation of vascular permeability (Hui et al., 2004; Moos et al., 2008), edema (Jiang et al., 2008), upregulation of cell adhesion molecules (Jiang et al., 2008; Ni et al., 2011) and myocardial infarction damage (Jiang et al., 2008; Ni et al., 2011). The ability of CysLT 2 R pharmacological antagonism to abolish these phenomena (Moos et al., 2008; Ni et al., 2011) lent support to the idea that endothelial-expressed CysLT 2 R was the principal mediator of these effects. Given the disparity in levels between endogenous murine CysLT 2 R expression and the overexpressed endothelial-targeted hCysLT 2 R transgene (Hui et al., 2004), it was also logical to believe that these biological responses were endothelium-driven. However, our experiments utilizing a novel Tie2-expressing cell-restricted CysLT 2 R overexpressing mouse model indicate that CysLT 2 R-mediated exacerbation of inflammation is only partially driven by endothelial cell-expressed receptors, and that other CysLT 2 R receptor niches may play a vital role in initiating and mediating the inflammatory response. Despite possessing the same overexpression of endothelial hCysLT 2 R as the EC model, EC/KO mice suffer significantly less severe myocardial damage following ischemia/reperfusion. As transgenic CysLT 2 Rs account for the vast majority of endothelial CysLT 2 R in EC mice (Hui et 128 al., 2004), it was not expected that removal of endogenous murine endothelial CysLT 2 R would affect endothelial CysLT 2 R-mediated mechanisms. Thus, our findings indicate that the endothelial CysLT 2 R population is not the only mediator of increased I/R injury in EC mice. We also note for the first time that KO mice show a tendency for reduced I/R damage versus WT counterparts. The reason that the difference between damage in KO and WT mice is relatively small compared to that between WT and EC mice is that basal CysLT 2 R expression in murine cardiac vasculature is quite low (Hui et al., 2001). However, basal human CysLT 2 R expression is higher (Heise et al., 2000), and leukotriene production and leukotriene receptor expression is elevated in pathological states (Carry et al., 1992; Jiang et al., 2008; Shi et al., 2012a), indicating that our EC model more closely represents human CysLT 2 R expression levels. Explaining our results required closer examination of the various pro-inflammatory mechanisms stemming from CysLT 2 R activation. We show here that basal rolling leukocyte flux is significantly higher in mice that possess endogenous “non-endothelial” CysLT 2 R (WT and EC groups) compared to mice that do not (KO and EC/KO groups), regardless of presence or absence of transgenic endothelial CysLT 2 R. While leukocyte recruitment to injured tissue is a key component of tissue repair (Jung et al., 2013; Kolaczkowska and Kubes, 2013), exacerbated leukocyte recruitment and improper resolution of the inflammatory response can result in additional injury (Kolaczkowska and Kubes, 2013). We have previously shown that transgenic overexpression of endothelial CysLT 2 R results in vascular hyperpermeability (Moos et al., 2008), and our EC/KO model was in agreement with this finding, showing significantly elevated extravascular FITC-albumin accumulation following cysLT stimulation. The extent and 129 magnitude of the vascular permeability response in EC/KO mice was comparable to that seen in EC mice, indicating that “non-endothelial” CysLT 2 R does not play a significant role in CysLT 2 R-mediated vascular hyperpermeability. The presence of rolling leukocytes does not automatically translate into leukocyte transmigration. Indeed, the presence of a patrolling vascular leukocyte reservoir consisting primarily of rolling or crawling monocytes that do not extravasate until following exposure to irritants and/or injury has been characterized in the mesenteric and coronary vasculature (Auffray et al., 2007; Jung et al., 2013). However, we saw a significant decrease in rolling leukocyte flux, as well as a transient, but significant, increase in slow-rolling leukocyte proportion in EC and EC/KO mice following cysLT stimulation. These findings, in combination with the aforementioned cysLT-induced hyperpermeability response in EC and EC/KO mice, support a mechanism where CysLT 2 R activation results in increased rolling leukocyte flux, decreased rolling leukocyte velocity, and vascular hyperpermeability, resulting in the elevated edema and leukocyte extravasation into the injured myocardium which have been shown to be hallmarks of exacerbated CysLT 2 R-mediated I/R injury (Jiang et al., 2008; Ni et al., 2011). However, while post-capillary cremaster venules in EC mice showed leakage in the absence of exogenous cysLT stimulation, vessels in EC/KO mice did not. Moos et al. (2008) attributed the phenomenon in EC mice to small amounts of leukotrienes liberated during the surgical exposure of the cremaster muscle taking advantage of an increased abundance of endothelial CysLT 2 R. If this is the case, why do these liberated leukotrienes not exert a similar effect on vessels in EC/KO mice? The answer may be that leukotrienes released during tissue injury do not directly 130 activate endothelial CysLT 2 R. Rather, trauma results in localized leukotriene production/release (Allen et al., 1993; Carry et al., 1992) and the subsequent activation of CysLT 2 Rs in sites other than the endothelium. This in turn results in cysLT production in close proximity to endothelial cells – a vital feature given the short half-lives of LTC 4 and LTD 4 – resulting in the activation of endothelial CysLT 2 R and autocrine cysLT production mechanisms. Sufficient leukotriene presence (i.e. exogenous application) would be able to bypass the “non-endothelial” step and directly activate endothelial receptors - explaining how EC/KO mice can still present hyperpermeability after exogenous stimulation. The exact cellular identity of these “nonendothelial” CysLT 2 R niches being activated in mice has yet to be determined, but leukocytes and other circulatory cells are leading candidates as CysLT 2 R is expressed by monocytes, macrophages, eosinophils (Heise et al., 2000), and platelets (Hasegawa et al., 2010) in humans. Eosinophils (Carnini et al., 2011), mast cells, macrophages (Funk, 2001), and parenchymal cells (Zarini et al., 2009) are capable of producing cysLTs, and neutrophils and monocytes produce the cysLT precursor LTA 4 (Di Gennaro et al., 2004; Folco and Murphy, 2006), which is converted to LTC 4 by endothelial cells and/or platelets (Folco and Murphy, 2006). Endothelial cells can produce LTC 4 in an autocrine manner upon CysLT 2 R activation in the presence of LTA 4 (Carnini et al., 2011). A limitation of the study is using the cremaster vasculature to extrapolate findings to the coronary circulation, given the known physiological and anatomical heterogeneity amongst microvascular beds (Aird, 2007). However, the observations in the cremaster vasculature do not contradict the post-I/R injury damage phenotypes detected in the myocardium. A novel and nonobtrusive method of real-time cardiac intravital microscopy imaging has just been realized (Jung 131 et al., 2013), affording an opportunity to examine the similarities and differences in response to cysLT-stimulation directly when this technique becomes more widely available. Furthermore, while the Tie2 promoter, used in our transgenic mice, specifies endothelial-targeted gene expression, it may also lead to expression in a small subset of monocytes, termed Tie2expressing monocytes (TEMs) (De Palma et al., 2005). These monocytes are highly angiogenic and are vital to tumor neovascularization, but also are mobilized following ischemia for the revascularization of ischemic tissue (Patel et al., 2013). Indeed, the majority of Tie2+ monocytes in mice are "resident" CD11b+Gr-1low/neg monocytes (Venneri et al., 2007), thought to be instrumental to inflammatory response resolution (Nahrendorf et al., 2010), so it will be interesting to investigate the role of CysLT 2 R activation/overexpression in this cell population in future studies. However, we do not propose that Tie2+ myeloid cells play a large role in the mechanisms explored herein, because overexpression of CysLT 2 R on Tie2+ myeloid cells would be common to both EC and ECKO mice, and thus would not explain the differences presented between the two genotypes. 4.5.2. Various tissue-specific CysLT 2 R niches play divergent, yet synergistic, roles in mediating exacerbation of CysLT 2 R-mediated ischemia/reperfusion injury. We previously proposed a mechanism where CysLT 2 R activation following ischemia/reperfusion injury resulted in elevated leukocyte extravasation and increased inflammation, resulting in increased tissue injury (Ni et al., 2011). Our present findings indicate that leukotriene release following injury/ischemia activates CysLT 2 R niches – likely circulating leukocytes – and that this facilitates leukocyte recruitment to the site of injury and activation of 132 endothelial CysLT 2 R (possibly via localized cysLT production). Endothelial CysLT 2 R activation results in vascular hyperpermeability – leading to edema – and an increase in the proportion of slow-rolling leukocytes, promoting leukocyte extravasation. Activation of both endothelial and “non-endothelial” CysLT 2 Rs likely yields a synergistic effect. Insufficient endothelial CysLT 2 R activation would result in higher numbers of rolling leukocytes that would not extravasate en masse (observed in WT mice), whereas absence of activation in locations other than the endothelium would result in limited rolling leukocyte flux (observed in EC/KO mice), and thus decreased leukocyte migration to the surrounding tissue regardless of vascular permeability level (Figure 4-8). In conclusion, we have used a novel endothelium-limited CysLT 2 R overexpressing mouse model to delineate the differing roles that endothelial and “non-endothelial” CysLT 2 R niches play in mediating post-injury inflammatory responses. We show that “non-endothelial” CysLT 2 R niches are vital to leukocyte recruitment and endothelial CysLT 2 R activation, whereas endothelial CysLT 2 R activation mediates extent of vascular permeability. While further work is required to clarify the precise molecular mechanisms underlying these phenotypes (i.e. cytokine release profile or induction of angiogenesis), as well as any involvement by other cell types (i.e. adipose tissue (Horrillo et al., 2010)) in close proximity to the vasculature, the revelation that endothelial CysLT 2 R activation alone does not govern CysLT 2 R-mediated pro-inflammatory responses indicates that more attention should be devoted to the role of CysLT 2 R expression in sites other than the endothelium in cardiovascular injury. 133 Figure 4-8: Proposed schematic for CysLT 2 R inflammatory response modulation. Injury or pro-inflammatory stimuli results in the synthesis and release of leukotrienes, which in turn activate CysLT 2 Rs present on resident and/or circulating cells – most likely leukocytes such as monocytes, macrophages, mast cells, and/or neutrophils. Activation of these CysLT 2 Rs results in increased leukocyte rolling flux, and promotes localized cysLT production, likely via supplying LTA 4 to endothelial cells and/or platelets. This results in activation of endothelial CysLT 2 Rs, causing increased slow rolling leukocyte fractions and vascular hyperpermeability – the latter contributing to edema in the affected tissue. These factors, in combination with the aforementioned increased leukocyte rolling flux, promote leukocyte transmigration into the surrounding tissue, resulting in exacerbation of the inflammatory response and elevated tissue damage. 134 4.6. Acknowledgements The authors would like to thank Professor Satoshi Ishii for providing LacZ-CysLT 2 R KO mice and Ms. Deborah Harrington for assistance with murine blood counts. This work was funded by the Canadian Institute for Health Research MOP-68930. C.D.F. holds a Tier 1 Canada Research Chair and is a Career Investigator of the Heart and Stroke Foundation of Ontario (Canada). 135 Chapter 5: General Discussion Although leukotrienes were first identified for their ability to induce smooth muscle contraction, research has since revealed that they are powerful inflammatory mediators as well. Much research has centered around CysLT 1 R-mediated effects in asthma and other respiratory tract diseases, and it was only recently that the role of cysteinyl leukotriene receptors in other inflammatory pathologies, especially cardiovascular disease, became a subject of discussion. This has brought the CysLT 2 R more into the spotlight, as it is the dominant isoform in the myocardium and coronary vasculature. However, gathering insight on pathways and mechanisms mediated by CysLT 2 R activation has been difficult due to a lack of CysLT 2 Rselective pharmacological agents in the past. In addition, while mouse models possessing altered CysLT 2 R expression levels do exist, drawing direct comparisons between endogenous human and murine CysLT 2 R expression profiles on a cellular level is also difficult due to a lack of murine-selective CysLT 2 R antibodies, resulting in an inability to localize CysLT 2 R expression to specific cell types. In an attempt to improve these situations, we have characterized a novel CysLT 2 R-selective antagonist both in vitro and in vivo, and used it to determine that CysLT 2 R is the sole cysteinyl leukotriene receptor mediating post-myocardial infarction responses in a murine model of CysLT 2 R overexpression. We have determined that exacerbation of CysLT 2 Rmediated myocardial infarction damage is due to enhanced activation of the postischemia/reperfusion inflammatory response, and that CysLT 2 R plays a minimal role during the ischemic phase or at the point of reperfusion. We have also uncovered that CysLT 2 Rs in different locations have varying roles in CysLT 2 R-mediated leukocyte recruitment and vascular hyperpermeability, and that “non-endothelial” CysLT 2 R activation may be required to trigger the hyperpermeability response via activation of endothelial receptors. Finally, we have explored 136 the effects of CysLT 2 R overexpression and CysLT 2 R blockade on long-term cardiac remodeling and heart function post-myocardial infarction. 5.1. Characterization of a Novel CysLT 2 R-Selective Antagonist Prior to 2010, the pharmacological agent most utilized in CysLT 2 R investigations was Bayu9773, a LTE 4 analogue that antagonized LTC 4 and LTD 4 -stimulated smooth muscle contraction in a dose-dependent manner (Tudhope et al., 1994). Bay-u9773 has equal affinity for CysLT 1 R and CysLT 2 R (Tudhope et al., 1994), but was also identified as a partial agonist for CysLT 2 R (Nothacker et al., 2000). Despite these drawbacks, Bay-u9773 remained the main pharmacological agent used in CysLT 2 R antagonism experiments until Wunder et al. (2010) published the first report of a CysLT 2 R-selective antagonist, dubbed HAMI3379. We have followed their work by characterizing the potency of a sister compound, termed BayCysLT 2 , in both cellular systems and murine models of pathology. HAMI3379 and BayCysLT 2 differ in molecular structure by one double bond (a cyclohexyl radical for the former, benzyl for the latter). Both are far more selective for CysLT 2 R versus CysLT 1 R, with HAMI3379 showing more than 10,000-fold selectivity for CysLT 2 R (Wunder et al., 2010) and BayCysLT 2 being at least 500-fold more selective for CysLT 2 R. In addition, HAMI3379 was found to be ~550-times more potent than the dual antagonist/partial agonist Bay-u9773 in blocking LTD 4 -mediated CysLT 2 R activation (Wunder et al., 2010). Unlike Bay-u9773, neither HAMI3379 nor BayCysLT 2 have partial agonist properties, determined by the lack of calcium release in CysLT 2 R or CysLT 1 R transfected cells during antagonist incubation periods (Wunder et al., 2010). IC 50 values determined from in vitro assays indicate that HAMI3379 is ~10-fold more 137 potent than BayCysLT 2 . However, experiments conducted with HAMI3379 by our laboratory have not corroborated this claim (Nassiri et al., unpublished data). One reason for this may be that we are using racemic mixtures of HAMI3379, whereas Wunder et al. may have experimented only with selected HAMI3379 stereoisomers. This would indicate that different stereoisomers of HAMI3379 have differing affinities for the CysLT 2 R, and also gives precedence that a similar phenomenon may apply to BayCysLT 2 as well. Additional experimentation is required to examine whether selected BayCysLT 2 stereoisomers in isolation have greater affinity than the racemic mixture used here. These discrepancies are unlikely to become a greater problem though, as both BayCysLT 2 and HAMI3379 are now commercially available, which standardizes the properties of the compound used by different research groups and prevents introduction of an additional variable. Our in vivo experimentation showed that BayCysLT 2 treatment abrogated exacerbation of CysLT 2 R overexpression-mediated myocardial infarction damage. However, more work is required in order to determine the pharmacokinetics and pharmacotoxicity of the compound. Carnini et al. (2011) have confirmed our findings that BayCysLT 2 is selective for CysLT 2 R in vitro, while Huang et al. (2008) and Qi et al. (2011) demonstrated BayCysLT 2 activity in cultured cells, but no other study to date has shed any light on the bioavailability, metabolism, or excretion of the compound. Similarly, HAMI3397 has been utilized in several studies since it was first characterized in vitro (Wunder et al., 2010), and its efficacy in cultured cells and in vivo is well established (Shi et al., 2012a; Zhang et al., 2013). However, as with BayCysLT 2 , little information is available regarding the pharmacokinetics and pharmacotoxicity of HAMI3379. Investigation of the pharmacological properties of BayCysLT 2 and HAMI3379 will be integral 138 to regulation of CysLT 2 R activity during pathological processes to identify when CysLT 2 R signaling is most critical. 5.2. Identification of CysLT 2 R-Mediated Mechanisms Post-Myocardial Infarction 5.2.1. CysLT 2 R is the main cysteinyl leukotriene receptor involved in myocardial infarction responses Jiang et al. (2008) have previously established that endothelial-targeted CysLT 2 R overexpression results in exacerbated myocardial damage following ischemia/reperfusion. However, the underlying mechanisms of this phenomenon remained unclear. At the time, the only CysLT 2 R antagonist available was the non-selective dual cysLTR antagonist Bay-u9773, which also had partial agonist properties against CysLT 2 R (Nothacker et al., 2000). While treatment with Bayu9773 attenuated exacerbation of myocardial infarction damage in EC mice, it also yielded ambiguity regarding whether CysLT 2 R was the sole mediator of the phenomenon, or whether CysLT 1 R was involved in some capacity. The synthesis of BayCysLT 2 has allowed us to isolate the role of CysLT 2 R in myocardial infarction damage exacerbation, and our results indicate that the phenomenon observed by Jiang et al. (2008) in EC mice is mainly mediated by CysLT 2 R, and that other leukotriene receptors play minimal roles (Ni et al., 2011). This finding fits given that CysLT 2 R is by far the dominant leukotriene receptor in human myocardium and endothelial cells (Heise et al., 2000; Lotzer et al., 2003). 139 One question that was raised (Moos and Funk, 2008) following the initial investigation by Jiang et al. (2008), and remains, is why does CysLT 2 R antagonism not reduce infarction size in wildtype mice? On the same note, why do CysLT 2 R KO mice not present significantly decreased infarct size compared to wild-type counterparts as well? While preliminary investigations by Jiang et al. (2008) indicated that KO mice did not show any reduction in infarction size versus wild-type counterparts, more in-depth study detailed here indicates that KO mice do indeed show slightly (but statistically non-significant) diminished myocardial damage following ischemia/reperfusion injury. Nonetheless, how CysLT 2 R expression affects infarction size presents a distinctly different pattern than how CysLT 2 R expression affects vascular permeability, where both receptor antagonism and genetic inactivation results in significantly decreased vascular permeability (Moos et al., 2008). One potential explanation is that endogenous CysLT 2 R expression in the heart and coronary circulation is very low. In contrast to the expression profile in humans (Heise et al., 2000), Hui et al. (2001) could not detect cardiac CysLT 2 R expression using Northern blot, and only identified endothelial CysLT 2 R expression on coronary vessels using the more sensitive in situ hybridization technique. As such, CysLT 2 R makes minimal or insignificant contributions to ischemia/reperfusion injury in mice under endogenous conditions. However, this does not mean that the findings observed in EC mice are merely the products of a contrived system. Even in mice, CysLT 2 R is physiologically upregulated in vasculature following ischemic stress: this has been observed in the heart and retina (Barajas-Espinosa et al., 2012; Jiang et al., 2008). Furthermore, leukotriene receptors are upregulated in pathological conditions, including atherosclerosis and cancer (Allen et al., 1998; Magnusson et al., 2007). As such, the EC mouse model is physiologically relevant, and is 140 important for the study of the effects of overstimulation of the post-infarction inflammatory response. 5.2.2. CysLT 2 R activation results in promotion of cell adhesion and vascular hyperpermeability Although we have identified the CysLT 2 R as the principle mediator of exacerbated myocardial infarction damage in EC mice, the underlying mechanisms triggered by CysLT 2 R activation remain unclear. We have pinpointed the CysLT 2 R as a mediator of post-ischemia/reperfusion inflammatory responses, based on the ability of CysLT 2 R-selective antagonist treatment to abrogate exacerbation of myocardial infarction injury even when delivered 45 minutes following reperfusion of the myocardium - a timepoint too late for administration of BayCysLT 2 to affect pathways triggered by ischemia, or prime the heart against acute reperfusion injury. Previous research shows that EC mice present increased myocardial leukocyte counts, as well as elevated red blood cell extravasation and myocardial edema following ischemia/reperfusion injury. Indeed, the ability of CysLT 2 R activation via exogenous leukotriene application to induce potent vascular leakage and edema is well known, and this response is also exacerbated in CysLT 2 R overexpressing mice (Hui et al., 2004; Moos et al., 2008). We confirm the finding of Jiang et al. by presenting evidence of elevated neutrophil extravasation into the myocardium following infarction (Ni et al., 2011). Furthermore, we show that CysLT 2 R antagonism can attenuate this effect, indicating that the leukocyte extravasation observed previously and the neutrophil presence presented here are likely CysLT 2 R-activation mediated. In addition to this, previous research shows that the genes encoding the cell adhesion molecules VCAM-1 and ICAM are upregulated following myocardial infarction, and that this upregulation is significantly elevated 141 in EC mice (Jiang et al., 2008). Our work here shows that these alterations in gene regulation are also attenuated via CysLT 2 R blockade, indicating that CysLT 2 R activation not only facilitates extravasation via induction of vascular permeability, but also by promoting leukocyte rolling and adhesion as well. These findings in the pathological heart are supported by investigations of CysLT 2 R-mediated phenomena in the peripheral cremaster microvasculature. While the cremaster vascular bed is not a perfect mimic of the coronary vasculature, it (or the similarly dissimilar mesenteric vascular bed) remains the main readily accessible site for real-time observation of vascular responses in vivo. Recent work has demonstrated that real-time intravital microscopy can be performed on a rapidly mobile tissue such as the heart, but the technology and apparatus required is not sufficiently available or affordable (Jung et al., 2013; Li et al., 2012). Moos et al. (2008) noted that cremaster post-capillary venule permeability was significantly reduced in CysLT 2 R KO mice and significantly elevated in EC mice. We have confirmed these findings and report that leukotriene stimulation of post-capillary venules results in increased leukocyte rolling flux over time, as well as an elevated proportion of E-selectin-mediated slow-rolling (Kunkel and Ley, 1996) leukocytes in wild-type mice. Neither of these phenomena was observed in CysLT 2 R KO mice, indicating that CysLT 2 R activation results in promotion of leukocyte recruitment and transmigration. Interestingly, EC mice showed steadily decreasing leukocyte rolling flux and a transient increase in the proportion of slow-rolling leukocytes. These results can likely be explained as a consequence of the aforementioned CysLT 2 R-mediated vascular hyperpermeability response facilitating elevated leukocyte transmigration - the number of cells leaving the vasculature exceeds the number being recruited, resulting in a net decrease in 142 vascular leukocyte flux and fewer slow-rolling (as opposed to fast-rolling or transmigrated) leukocytes. Further investigation is required to confirm that leukocyte diapedesis and transmigration is occurring following leukotriene stimulation, but these findings provide rationale for the elevated leukocyte presence observed in the myocardium following ischemia/reperfusion injury in EC mice. 5.2.3. CysLT 2 R activation on different cell types mediate distinct and separate responses While we were able to establish several pro-inflammatory pathways triggered by CysLT 2 R activation post-ischemia/reperfusion, we were unable to isolate the specific role played by receptors present on endothelial cells versus those on other cell types. To investigate this, we crossed CysLT 2 R KO mice with endothelial CysLT 2 R overexpressing mice, yielding a mouse model (EC/KO) that does not express endogenous murine CysLT 2 R, but does expresses the human CysLT 2 R in the vascular endothelium (as well as a small subset of Tie2+ leukocytes) in comparable abundance to EC mice. Transgenic CysLT 2 R expression is significantly higher than endogenous CysLT 2 R expression in EC mice (Hui et al., 2004), and as a result, we anticipated that removing endogenous murine endothelial receptors would not impair any endothelial cell CysLT 2 R-mediated response. In addition, because the responses (i.e. vascular permeability and ICAM-1/VCAM expression) outlined in the previous section were largely endothelium mediated, we hypothesized that removing non-endothelial endogenous CysLT 2 R would not alter the myocardial infarction injury exacerbation phenomenon observed in EC mice. Surprisingly, EC/KO mice did not show exacerbated myocardial damage following ischemia/reperfusion 143 injury, indicating that the endothelial CysLT 2 R population was not the sole mediator of CysLT 2 R-mediated injury exacerbation. Further investigation of EC/KO mice showed that this genotype differed from EC mice in several ways. First, while cremaster muscle post-capillary venule hyperpermeability was present in EC/KO mice, it was only present when the muscle vasculature was stimulated with exogenous cysLTs. This is in contrast to EC mice, which show significantly elevated vascular permeability post-surgery prior to exogenous cysLT application. This phenomenon was attributed to CysLT 2 R activation by leukotrienes released from the tissue during the surgical preparation of the cremaster muscle for imaging (Moos et al., 2008). However, the finding that this can be abolished in EC/KO mice indicates that if there are leukotrienes being released during the surgical preparation of the cremaster muscle, they are not directly activating endothelial CysLT 2 R. Rather, it is perhaps more plausible that physical trauma to the cremaster muscle during surgical preparation releases leukotrienes, which may activate circulating leukocytes via CysLT 2 R binding, in turn resulting in further leukotriene production either by or in close proximity to endothelial cells. There is a lack of evidence indicating that cysLT receptor activation directly results in leukotriene formation, but both CysLT 1 R and CysLT 2 R activation results in increased intracellular Ca2+ concentrations (Lotzer et al., 2003; Lynch et al., 1999), which may serve to promote cPLAα2 and 5-LO translocation (Glover et al., 1995; Rouzer et al., 1985). In addition, there is ample evidence suggesting cysLT receptor activation promotes leukocyte recruitment and adhesion (Datta et al., 1995; Fregonese et al., 2002; McIntyre et al., 1986; Pedersen et al., 1997). This in turn may promote transcellular production of cysteinyl 144 leukotrienes, as leukocytes, which possess LTA 4 H, are brought into close proximity to endothelial cells and platelets, which possess LTC 4 S (Zarini et al., 2009). An alternative possibility is that removal of non-endothelial CysLT 2 R has increased the activation threshold for endothelial CysLT 2 R in some manner, and the limited amount of LTs released by surgical trauma can no longer activate endothelial receptors. CysLT receptors, like other GPCRs, do form hetero- and homodimers for the purposes of self-regulation. Indeed, in CysLT 1 R/CysLT 2 R heterodimers, activation of CysLT 2 R serves as a negative regulator of CysLT 1 R-mediated pathways (Jiang et al., 2007). CysLT receptors can also form heterodimers with GPR17 (Maekawa et al., 2009), and there is putative evidence for cross-talk between the classical cysLT receptors and the newly identified CysLT E R (GPR99) (Kanaoka et al., 2013). However, there are no reports thus far of cysLT receptor conformation and/or ligand affinities being altered via cell-cell interactions as opposed to cross-talk between receptors located on the same cell, and there is little evidence to support that cell-cell cysLT receptor modulation takes place. We also noted that leukocyte rolling flux counts under basal conditions are significantly diminished in mice lacking non-endothelial CysLT 2 R (KO and EC/KO genotypes) compared to mice that do express endogenous receptors (WT and EC genotypes). No significant differences were noted between WT and EC or KO and EC/KO groups, indicating that the presence/absence of endothelial CysLT 2 R overexpression did not play a role in this phenomenon. Furthermore, rolling leukocyte median velocity at 15 minutes post-LT stimulation was also significantly faster in KO and EC/KO mice versus WT and EC counterparts. These findings indicate that activation 145 of non-endothelial CysLT 2 Rs plays a role in facilitating and/or promoting leukocyte rolling under both endogenous and pro-inflammatory conditions. It is unclear how CysLT 2 R activation is accomplishing this, but cysLTs are well established as mediators of leukocyte chemotaxis (Fregonese et al., 2002; Hashimoto et al., 2009; Ichiyama et al., 2005) and promoters of leukocyte rolling/adhesion (Datta et al., 1995; McIntyre et al., 1986). There is ample evidence that these phenomena can be regulated by CysLT 1 R activation (Back et al., 2011; Boehmler et al., 2009), but that does not rule out a role for CysLT 2 R. Certainly, the lack of a selective CysLT 2 R antagonist until recently (Wunder et al., 2010) likely contributes to the disparity in information available about CysLT 1 R versus CysLT 2 R in mediating cysteinyl leukotrienetriggered chemotactic responses. Although experimental data from the EC/KO mouse model revealed that endothelial CysLT 2 R activation did not play as significant a role as previously anticipated, the receptor niche is still a vital component in exacerbation of CysLT 2 R-mediated damage. EC/KO mice are still capable of presenting significantly elevated vascular permeability upon leukotriene stimulation - indeed, vascular leakage seen in EC/KO mice is on par with EC counterparts and significantly higher than that observed in wild-type mice. We also observed that mice possessing endothelial CysLT 2 R overexpression (both EC and EC/KO groups) showed a decline in rolling leukocyte flux following LT stimulation. Finally, EC and EC/KO mice both show a significant increase in slow-rolling leukocyte proportion following leukotriene stimulation. In combination, these results indicate that endothelial CysLT 2 R activation mediates vascular hyperpermeability in tandem with promoting E-selectin mediated leukocyte slow-rolling. These two factors most likely facilitate increased leukocyte transmigration into the extravascular tissue, and this provides 146 an explanation for the steady decrease in rolling leukocyte flux post-LT stimulation observed in mice possessing elevated endothelial CysLT 2 R expression. The steady decrease in rolling leukocyte flux also provides an explanation for the transient nature of the increase in slow-rolling leukocyte proportion observed in EC and EC/KO mice. The extravasating cells would largely be drawn from the slow-rolling cell population, thus reducing the proportion of slow-rolling leukocytes present. Vascular hyperpermeability, leukocyte recruitment, and facilitation of leukocyte-slow rolling combine to create a scenario that greatly promotes and facilitates leukocyte extravasation to the extravascular milieu. This elevated leukocyte presence in the injured tissue would then lead to exacerbation of pro-inflammatory responses, leading to increased tissue damage and death. Indeed, neutrophil depletion (Romson et al., 1983), as well as genetic knockout of P-selectin and ICAM-1 (Briaud et al., 2001) has been linked to decreased acute myocardial injury. Furthermore, neutrophil depletion and inhibition of neutrophil adherence to endothelium both resulted in attenuation of I/R-induced vascular permeability (Hernandez et al., 1987). Based on this model, CysLT 2 Rs located on the endothelium and elsewhere both play important roles in exacerbation of CysLT 2 R-mediated injury. If “non-endothelial” receptors are not activated, then fewer rolling leukocytes are present in the circulation, resulting in lower extravasation despite the presence of a hyperpermeability response (as observed in EC/KO) mice. If endothelial CysLT 2 Rs are not expressed in sufficient quantities, then activation fails to yield a hyperpermeability response, resulting in increased vascular leukocyte rolling flux, but lower extravasation as the cells are unable to transmigrate (as observed in WT mice). If neither 147 CysLT 2 R subset is activated, then myocardial infarction damage is reduced (as seen in KO mice). Only when both receptor subsets are activated - and endothelial CysLT 2 R is present in significant quantities - do we observe exacerbation of injury (as seen in EC mice). 5.2.4. The CysLT 2 R picture is far from complete Several outstanding questions do remain however within this framework. First, what is the source of leukotrienes for CysLT 2 R activation in either leukocytes or endothelial cells? There is evidence for leukotriene release/production during ischemia/reperfusion injury (Carry et al., 1992; Takase et al., 1996). However, there is little information regarding the where and the when. Where are leukotrienes being released - in close enough proximity to relevant cells in order to exert their effects prior to conversion to LTE 4 ? When are they being released? Our experiments using BayCysLT 2 indicate that CysLT 2 R activation occurs predominantly postreperfusion, but the precise timeframe of LT release during and after MI remains unclear. It is also important to investigate cell-cell interactions that may arise prior to or upon CysLT 2 R activation. Given the prevalence of transcellular LT production, are certain cell-cell interactions (i.e. neutrophil-endothelial cell) required for the generation of sufficient quantities of LTs to activate CysLT 2 Rs? Conversely, could initial CysLT 2 R activation promote cell-cell interactions, resulting in additional local LT production and enhanced CysLT 2 R activation? Second, aside from endothelial cells, where is CysLT 2 R endogenously expressed in the murine model? In humans, CysLT 2 R is expressed by macrophages, eosinophils, mast cells, monocytes, and platelets (Hasegawa et al., 2010; Heise et al., 2000; Mellor et al., 2003; Mita et al., 2001), 148 but as previously mentioned, cell-specific localization has been difficult due to absence of a commercially available murine-specific antibody, as well as lower endogenous receptor expression (Hui et al., 2001). We generate our hypotheses and models working under the assumption that murine and human CysLT 2 R expression profiles are relatively similar. Data from when the human and murine receptor sequences were first identified indicate that this is the case, at least at the organ-level. Both human and mouse showed strongest CysLT 2 R expression in myeloid cell containing tissues (i.e. spleen, thymus, and lymph nodes), and found lesser expression in peripheral blood leukocytes, brain, and heart. The main difference though was that relative expression of CysLT 2 R in the mouse brain, heart, and leukocytes was significantly lower versus organs of high expression (i.e. spleen) compared to human equivalents (Heise et al., 2000; Hui et al., 2001). Assuming that mice, like humans, express CysLT 2 R on circulating leukocytes, one question is whether CysLT 2 R is expressed on pro-inflammatory leukocytes, or whether the receptor is present on anti-inflammatory cells, such as M2 macrophages, Ly-6Clo monocytes, or regulatory T cells (Hofmann et al., 2012; Nahrendorf et al., 2007). Aside from circulating leukocytes, another cell type that is a candidate for CysLT 2 R expression and can modulate endothelial function is the pericyte. Pericytes are mural cells that share a basement membrane with endothelial cells and are vital to the regulation of vascular function (Armulik et al., 2005). They can interact directly with multiple endothelial cells simultaneously and are present in the microvasculature, as well as large arteries and veins. Pericyte activation regulates vasoconstriction and vasodilation in vascular beds, and play a critical role in maintenance of the blood-brain barrier (Bergers and Song, 2005). Barajas-Espinosa et al. (2012) reported CysLT 2 R expression on vascular pericytes in the retina, and given their role in regulation of vascular 149 function, it is possible that they may play a role in mediating leukotriene-induced hyperpermeability responses. Third, how does CysLT 2 R activation on peripheral cells result in endothelial CysLT 2 R activation? The most likely hypothesis is that CysLT 2 R activation results in leukotriene production by circulating leukocytes. Whether this occurs due to direct targeting of leukotriene production pathway enzymes or occurs indirectly as a by-product of a combination of increased intracellular Ca2+ and the formation of a pro-inflammatory environment is unknown. The leukotriene produced would likely be LTA 4 , which would then lead to cysLT production via transcellular biosynthesis. As outlined in the introduction, cysLTs can be produced by platelets and endothelial cells in humans using donor LTA 4 from other cell types (Maclouf and Murphy, 1988; Sjostrom et al., 2001). This phenomenon has also been confirmed in murine models, although the exact donor and production cell types are unclear (Zarini et al., 2009). There is evidence that cysteinyl leukotrienes can exert autocrine effects in endothelial cells (Carnini et al., 2011), fibrocytes (Vannella et al., 2007), and eosinophils (Lee et al., 2000), so it is possible that endothelial cell LT production in our model exacerbates vascular permeability through autocrine activation of elevated numbers of endothelial CysLT 2 Rs. However, further work is certainly required to pinpoint the specific details of the in vivo mechanism upon ischemic injury. Finally, is CysLT 2 R activation always deleterious – i.e. will inactivation of CysLT 2 R signaling as part of a course of CVD treatment prove to be ultimately detrimental? The large body of work investigating the pathological implications of CysLT 2 R activation far overshadows the number of investigations regarding the normal endogenous function of this receptor. Studies show that 150 IL-4 plays a role in promoting CysLT 2 R activation, either via receptor upregulation (Lotzer et al., 2003) or by priming cells expressing CysLT 2 R (Mellor et al., 2003). This may indicate that CysLT 2 R activation is part of host defense, as IL-4 facilitates Th-2 effector cell-mediated counter-mechanisms against parasitic worm invasion (Chen et al., 2004a). Certainly, as mentioned in the introductory chapter of this manuscript, CysLT 2 R activation has been shown to facilitate upregulation and secretion of IL-8, a potent stimulator of neutrophil chemotaxis (Thompson et al., 2008). The fact that CysLT 2 R expression in HUVECs is rapidly downregulated by pro-inflammatory stimuli (i.e. LPS, TNF-α) (Sjostrom et al., 2003) indicates that CysLT 2 R-mediated inflammatory mechanisms are designed to be transient, and that pathology likely arises from uncontrolled increases in either stimuli or an inability to shut off inflammatory mechanisms. Research has also shown that CysLT 2 R activation promotes cell proliferation (Huang et al., 2008; Vannella et al., 2007). Conversely, there is also evidence that CysLT 2 R may suppress cell proliferation. High CysLT 2 R expression, in conjunction with low CysLT 1 R expression, has been associated with decreased disease severity and improved prognosis in colorectal cancer patients, indicating that in this situation, CysLT 2 R signaling may be impeding CysLT 1 R-mediated cell survival/proliferation pathways (Magnusson et al., 2007). However, these studies were conducted in pathological models, which do not clarify the role of CysLT 2 R in mediating cell proliferation/differentiation under basal conditions, but do imply that CysLT 2 R activation may regulate the cell cycle. There is also the possibility that CysLT 2 R may also serve as a negative regulator of CysLT 1 R or CysLT E R. Indeed, aside from the above mentioned example in colorectal cancer cells, CysLT 2 R knockdown results in increased CysLT 1 R-mediated mast cell proliferation and the enhancement of cysLT-mediated ERK phosphorylation (Jiang et al., 2007). Furthermore, knockout of CysLT 1 R and CysLT 2 R in mice 151 resulted in increased vascular ear edema in response to LTE 4 stimulation compared to wild-type animals, indicating that the presence of those two receptors served to dampen the proinflammatory activity of the CysLT E R (Kanaoka et al., 2013). Certainly, further investigation is warranted in comparing and contrasting CysLT 2 R-mediated responses in physiological and pathological conditions. 5.3. CysLT 2 R Antagonism Post-Reperfusion Impedes Long-Term Ventricular Remodeling While we have proposed a putative mechanism for exacerbation of CysLT 2 R-mediated acute myocardial injury, how CysLT 2 R activation affects long-term cardiac function postischemia/reperfusion injury remains unclear. We investigated long-term cardiac function indirectly in WT and EC mice following myocardial infarction and surprisingly observed no prominent differences between the two groups, despite the disparity in acute myocardial injury. Furthermore, and even more surprisingly, we found that BayCysLT 2 treatment in both WT and EC mice actually results in decreased cardiac function versus untreated counterparts, despite apparently limiting acute myocardial infarction damage based on the significantly greater apical region ventricular wall thickness observed in BayCysLT 2 treated mice. These findings are certainly puzzling, and yet perhaps not inherently contradictory. Little is known about the role of CysLT 2 R in inflammatory resolution, so it is possible that CysLT 2 R antagonism exerts both beneficial (prevention of elevated inflammatory response) and deleterious (inhibition of proper debris clearance and wound healing) effects on the infarcted heart. Certainly, any discrepancy in CysLT 2 R expression levels between pro-inflammatory (e.g. 152 Ly-6Chi monocytes and M1 macrophages) and anti-inflammatory leukocyte subsets (e.g. Ly-6Clo monocytes and M2 macrophages) is worthy of investigation. Furthermore, there is evidence that in certain systems, CysLT 2 R mediates beneficial pathways. For example, CysLT 2 R activation has been shown to negatively regulate CysLT 1 R-mediated responses in mast cells, and CysLT 2 R knockdown results in increased CysLT 1 R expression (Jiang et al., 2007). In addition, elevated CysLT 2 R expression has been associated with cell differentiation in colorectal cancer (Magnusson et al., 2007) and positive prognosis in breast cancer (Magnusson et al., 2011) and prostate cancer (Matsuyama et al., 2007). It must be noted that while these studies indicate a role for CysLT 2 R as a counter-balance to CysLT 1 R, we do not believe that CysLT 1 R plays a significant role in myocardial infarction injury, given the paucity of the receptor in the heart. However, these examples do serve to illustrate the possibility of a CysLT 2 R-mediated mechanism that is beneficial to cardiac function post-injury. Another possible explanation for our findings is that our transient ischemia/reperfusion may not sufficiently damage the myocardium to evoke noticeable cardiac remodeling in our model, and this is masking CysLT 2 R-mediated changes to long-term cardiac function. While the literature presents many examples of studies utilizing transient ischemia/reperfusion demonstrating significant differences between experimental groups (Arslan et al., 2013; Eguchi et al., 2012; Toldo et al., 2012), there is a study that rejected animals suffering under 30% infarction size under the rationale that they "did not show typical LV remodeling" (Harada et al., 1999). In addition, the literature available would be biased towards cases where I/R induced significant changes versus cases where no significant changes were noted, given the difficulty in publishing “negative results.” One possible remedy for this is to use a model of permanent LAD ligation in 153 order to induce ischemia - certainly, studies that utilize permanent ligation present much more pronounced wall thinning and ventricular dilatation than that observed in our investigation (Patten et al., 1998; Song et al., 2012; Zhang et al., 2005). In addition, studies (albeit those using permanent LAD ligation) have also noted dilatation in the right ventricle, as well as the left postMI, and that was something not investigated in the present study (Patten et al., 1998; Yang et al., 2002). However, this technique is not without its own drawbacks, as it ostensibly bypasses reperfusion injury and the subsequent inflammatory response. Certainly, the role of CysLT 2 R activation in long-term cardiac remodeling and function warrants further and more in-depth attention. 5.4. Potential for the CysLT 2 R as a Therapeutic Target While CysLT 1 R is well-known as a target in the treatment of asthma and other inflammatory airway diseases, CysLT 2 R has only recently gained attention as a potential therapeutic target in cardiovascular disease treatment. Although we have characterized a novel CysLT 2 R-selective antagonist, and firmly established a role for CysLT 2 R activation-mediated mechanisms in the exacerbation of acute myocardial infarction injury, much work remains to be done. No information is currently available regarding the pharmacological properties of BayCysLT 2 , and no side effects of systemic administration of the compound have been reported to this date. In addition, very little is known about the bioavailability or metabolism of HAMI3379 or BayCysLT 2 . Another important issue is that although murine models are highly useful due to the ability to manipulate genetic expression of targets of investigation, the endogenous CysLT 2 R expression profile in the murine cardiovascular system does not fully mimic that in humans. In 154 particular, low endogenous expression of CysLT 2 R in the heart and coronary vasculature makes it difficult to examine CysLT 2 R-specific effects and mechanisms without genetic manipulation. In conclusion, while CysLT 2 R has been investigated for almost 15 years now, our understanding of the receptor and its effects is very much still in its infancy. Understanding how leukotriene stimulation activates and regulates inflammatory responses post-injury is not only vital to improving patient outcome post-myocardial infarction, it may also provide a novel approach to treating inflammation, both chronic and acute, caused by other pathologies including atherosclerosis and stroke. Certainly, persistent cysLT receptor antagonism would be a novel way to treat deleterious gene upregulation present during chronic pathologies (i.e. proinflammatory genes upregulated by leukotriene stimulation (Back et al., 2011; Eaton et al., 2012; Uzonyi et al., 2006) or provide a counter-measure to upregulation of CysLT receptors present during atherosclerosis and stenosis (Allen et al., 1998; Nagy et al., 2011). While much more investigation into the precise mechanisms of CysLT 2 R activation post-myocardial infarction is clearly required, the advent of proper pharmacological tools capable of isolating CysLT 2 Rmediated effects makes clarifying the role of CysLT 2 R in cardiovascular pathogenesis much easier, and will hopefully lead to treatment strategies targeting cardiovascular disease and inflammation. 155 References Abbracchio, M.P., Burnstock, G., Boeynaems, J.M., Barnard, E.A., Boyer, J.L., Kennedy, C., Miras-Portugal, M.T., King, B.F., Gachet, C., Jacobson, K.A., et al. (2005). The recently deorphanized GPR80 (GPR99) proposed to be the P2Y15 receptor is not a genuine P2Y receptor. Trends in pharmacological sciences 26, 8-9. Abramovitz, M., Wong, E., Cox, M.E., Richardson, C.D., Li, C., and Vickers, P.J. (1993). 5lipoxygenase-activating protein stimulates the utilization of arachidonic acid by 5-lipoxygenase. European journal of biochemistry / FEBS 215, 105-111. Adelroth, E., Morris, M.M., Hargreave, F.E., and O'Byrne, P.M. (1986). Airway responsiveness to leukotrienes C4 and D4 and to methacholine in patients with asthma and normal controls. The New England journal of medicine 315, 480-484. Ahn, J., and Kim, J. (2012). Mechanisms and consequences of inflammatory signaling in the myocardium. Current hypertension reports 14, 510-516. Aird, W.C. (2007). Phenotypic heterogeneity of the endothelium: II. Representative vascular beds. Circulation research 100, 174-190. Albelda, S.M., Smith, C.W., and Ward, P.A. (1994). Adhesion molecules and inflammatory injury. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 8, 504-512. Allen, S., Dashwood, M., Morrison, K., and Yacoub, M. (1998). Differential leukotriene constrictor responses in human atherosclerotic coronary arteries. Circulation 97, 2406-2413. Allen, S.P., Sampson, A.P., Piper, P.J., Chester, A.H., Ohri, S.K., and Yacoub, M.H. (1993). Enhanced excretion of urinary leukotriene E4 in coronary artery disease and after coronary artery bypass surgery. Coronary artery disease 4, 899-904. Amat, M., Diaz, C., and Vila, L. (1998). Leukotriene A4 hydrolase and leukotriene C4 synthase activities in human chondrocytes: transcellular biosynthesis of Leukotrienes during granulocytechondrocyte interaction. Arthritis and rheumatism 41, 1645-1651. Amsterdam, E.A., Pan, H.L., Rendig, S.V., Symons, J.D., Fletcher, M.P., and Longhurst, J.C. (1993). Limitation of myocardial infarct size in pigs with a dual lipoxygenase-cyclooxygenase blocking agent by inhibition of neutrophil activity without reduction of neutrophil migration. Journal of the American College of Cardiology 22, 1738-1744. Armulik, A., Abramsson, A., and Betsholtz, C. (2005). Endothelial/pericyte interactions. Circulation research 97, 512-523. Arslan, F., Lai, R.C., Smeets, M.B., Akeroyd, L., Choo, A., Aguor, E.N., Timmers, L., van Rijen, H.V., Doevendans, P.A., Pasterkamp, G., et al. (2013). Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to 156 enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem cell research 10, 301-312. Auffray, C., Fogg, D., Garfa, M., Elain, G., Join-Lambert, O., Kayal, S., Sarnacki, S., Cumano, A., Lauvau, G., and Geissmann, F. (2007). Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 317, 666-670. Austen, K.F. (2005). The mast cell and the cysteinyl leukotrienes. Novartis Foundation symposium 271, 166-175; discussion 176-168, 198-169. Austen, K.F., Maekawa, A., Kanaoka, Y., and Boyce, J.A. (2009). The leukotriene E4 puzzle: finding the missing pieces and revealing the pathobiologic implications. The Journal of allergy and clinical immunology 124, 406-414; quiz 415-406. Back, M., Dahlen, S.E., Drazen, J.M., Evans, J.F., Serhan, C.N., Shimizu, T., Yokomizo, T., and Rovati, G.E. (2011). International Union of Basic and Clinical Pharmacology. LXXXIV: leukotriene receptor nomenclature, distribution, and pathophysiological functions. Pharmacological reviews 63, 539-584. Back, M., Qiu, H., Haeggstrom, J.Z., and Sakata, K. (2004). Leukotriene B4 is an indirectly acting vasoconstrictor in guinea pig aorta via an inducible type of BLT receptor. American journal of physiology Heart and circulatory physiology 287, H419-424. Bandeira-Melo, C., Hall, J.C., Penrose, J.F., and Weller, P.F. (2002). Cysteinyl leukotrienes induce IL-4 release from cord blood-derived human eosinophils. The Journal of allergy and clinical immunology 109, 975-979. Barajas-Espinosa, A., Ni, N.C., Yan, D., Zarini, S., Murphy, R.C., and Funk, C.D. (2012). The cysteinyl leukotriene 2 receptor mediates retinal edema and pathological neovascularization in a murine model of oxygen-induced retinopathy. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 26, 1100-1109. Barajas-Espinosa, A., Ochoa-Cortes, F., Moos, M.P., Ramirez, F.D., Vanner, S.J., and Funk, C.D. (2011). Characterization of the cysteinyl leukotriene 2 receptor in novel expression sites of the gastrointestinal tract. The American journal of pathology 178, 2682-2689. Barnes, N., Thomas, M., Price, D., and Tate, H. (2005). The national montelukast survey. The Journal of allergy and clinical immunology 115, 47-54. Barral, S., Fernandez-Cadenas, I., Bis, J.C., Montaner, J., Ikram, A.M., Launer, L.J., Fornage, M., Schmidt, H., Brickman, A.M., Seshadri, S., et al. (2012). No association of ALOX5AP polymorphisms with risk of MRI-defined brain infarcts. Neurobiology of aging 33, 629 e621623. Barry, M., Cahill, R.A., Roche-Nagle, G., Neilan, T.G., Treumann, A., Harmey, J.H., and Bouchier-Hayes, D.J. (2009). Neoplasms escape selective COX-2 inhibition in an animal model of breast cancer. Irish journal of medical science 178, 201-208. 157 Baud, L., Goetzl, E.J., and Koo, C.H. (1987). Stimulation by leukotriene D4 of increases in the cytosolic concentration of calcium in dimethylsulfoxide-differentiated HL-60 cells. The Journal of clinical investigation 80, 983-991. Bautz, F., Denzlinger, C., Kanz, L., and Mohle, R. (2001). Chemotaxis and transendothelial migration of CD34(+) hematopoietic progenitor cells induced by the inflammatory mediator leukotriene D4 are mediated by the 7-transmembrane receptor CysLT1. Blood 97, 3433-3440. Belhomme, D., Peynet, J., Louzy, M., Launay, J.M., Kitakaze, M., and Menasche, P. (1999). Evidence for preconditioning by isoflurane in coronary artery bypass graft surgery. Circulation 100, II340-344. Benned-Jensen, T., and Rosenkilde, M.M. (2010). Distinct expression and ligand-binding profiles of two constitutively active GPR17 splice variants. British journal of pharmacology 159, 1092-1105. Bergers, G., and Song, S. (2005). The role of pericytes in blood-vessel formation and maintenance. Neuro-oncology 7, 452-464. Bernstrom, K., and Hammarstrom, S. (1981). Metabolism of leukotriene D by porcine kidney. The Journal of biological chemistry 256, 9579-9582. Bevan, S., Dichgans, M., Wiechmann, H.E., Gschwendtner, A., Meitinger, T., and Markus, H.S. (2008). Genetic variation in members of the leukotriene biosynthesis pathway confer an increased risk of ischemic stroke: a replication study in two independent populations. Stroke; a journal of cerebral circulation 39, 1109-1114. Bilir, A., Erkasap, N., Koken, T., Gulec, S., Kaygisiz, Z., Tanriverdi, B., and Kurt, I. (2007). Effects of tramadol on myocardial ischemia-reperfusion injury. Scandinavian cardiovascular journal : SCJ 41, 242-247. Birdsall, H.H., Green, D.M., Trial, J., Youker, K.A., Burns, A.R., MacKay, C.R., LaRosa, G.J., Hawkins, H.K., Smith, C.W., Michael, L.H., et al. (1997). Complement C5a, TGF-beta 1, and MCP-1, in sequence, induce migration of monocytes into ischemic canine myocardium within the first one to five hours after reperfusion. Circulation 95, 684-692. Bisoendial, R.J., Tanck, M.W., Golledge, J., Broekhuizen, L.N., Legemate, D.A., Stroes, E.S., and Norman, P.E. (2012). The association between the gene encoding 5-lipoxygenase activating protein and abdominal aortic aneurysms. Atherosclerosis 220, 425-428. Boehmler, A.M., Drost, A., Jaggy, L., Seitz, G., Wiesner, T., Denzlinger, C., Kanz, L., and Mohle, R. (2009). The CysLT1 ligand leukotriene D4 supports alpha4beta1- and alpha5beta1mediated adhesion and proliferation of CD34+ hematopoietic progenitor cells. Journal of immunology 182, 6789-6798. Borgeat, P., Hamberg, M., and Samuelsson, B. (1976). Transformation of arachidonic acid and homo-gamma-linolenic acid by rabbit polymorphonuclear leukocytes. Monohydroxy acids from novel lipoxygenases. The Journal of biological chemistry 251, 7816-7820. 158 Borgeat, P., and Samuelsson, B. (1979a). Arachidonic acid metabolism in polymorphonuclear leukocytes: effects of ionophore A23187. Proceedings of the National Academy of Sciences of the United States of America 76, 2148-2152. Borgeat, P., and Samuelsson, B. (1979b). Arachidonic acid metabolism in polymorphonuclear leukocytes: unstable intermediate in formation of dihydroxy acids. Proceedings of the National Academy of Sciences of the United States of America 76, 3213-3217. Braccioni, F., Dorman, S.C., O'Byrne P, M., Inman, M.D., Denburg, J.A., Parameswaran, K., Baatjes, A.J., Foley, R., and Gauvreau, G.M. (2002). The effect of cysteinyl leukotrienes on growth of eosinophil progenitors from peripheral blood and bone marrow of atopic subjects. The Journal of allergy and clinical immunology 110, 96-101. Brach, M.A., de Vos, S., Arnold, C., Gruss, H.J., Mertelsmann, R., and Herrmann, F. (1992). Leukotriene B4 transcriptionally activates interleukin-6 expression involving NK-chi B and NFIL6. European journal of immunology 22, 2705-2711. Brady, H.R., and Serhan, C.N. (1992). Adhesion promotes transcellular leukotriene biosynthesis during neutrophil-glomerular endothelial cell interactions: inhibition by antibodies against CD18 and L-selectin. Biochemical and biophysical research communications 186, 1307-1314. Briaud, S.A., Ding, Z.M., Michael, L.H., Entman, M.L., Daniel, S., and Ballantyne, C.M. (2001). Leukocyte trafficking and myocardial reperfusion injury in ICAM-1/P-selectin-knockout mice. American journal of physiology Heart and circulatory physiology 280, H60-67. Brock, T.G., McNish, R.W., Bailie, M.B., and Peters-Golden, M. (1997). Rapid import of cytosolic 5-lipoxygenase into the nucleus of neutrophils after in vivo recruitment and in vitro adherence. The Journal of biological chemistry 272, 8276-8280. Brocklehurst, W.E. (1960). The release of histamine and formation of a slow-reacting substance (SRS-A) during anaphylactic shock. The Journal of physiology 151, 416-435. Buja, L.M. (2005). Myocardial ischemia and reperfusion injury. Cardiovascular pathology : the official journal of the Society for Cardiovascular Pathology 14, 170-175. Byrum, R.S., Goulet, J.L., Snouwaert, J.N., Griffiths, R.J., and Koller, B.H. (1999). Determination of the contribution of cysteinyl leukotrienes and leukotriene B4 in acute inflammatory responses using 5-lipoxygenase- and leukotriene A4 hydrolase-deficient mice. Journal of immunology 163, 6810-6819. Canetti, C., Hu, B., Curtis, J.L., and Peters-Golden, M. (2003). Syk activation is a leukotriene B4-regulated event involved in macrophage phagocytosis of IgG-coated targets but not apoptotic cells. Blood 102, 1877-1883. Capra, V., Ravasi, S., Accomazzo, M.R., Citro, S., Grimoldi, M., Abbracchio, M.P., and Rovati, G.E. (2005). CysLT1 receptor is a target for extracellular nucleotide-induced heterologous desensitization: a possible feedback mechanism in inflammation. Journal of cell science 118, 5625-5636. 159 Carden, D.L., and Granger, D.N. (2000). Pathophysiology of ischaemia-reperfusion injury. The Journal of pathology 190, 255-266. Carnini, C., Accomazzo, M.R., Borroni, E., Vitellaro-Zuccarello, L., Durand, T., Folco, G., Rovati, G.E., Capra, V., and Sala, A. (2011). Synthesis of cysteinyl leukotrienes in human endothelial cells: subcellular localization and autocrine signaling through the CysLT2 receptor. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 25, 3519-3528. Carry, M., Korley, V., Willerson, J.T., Weigelt, L., Ford-Hutchinson, A.W., and Tagari, P. (1992). Increased urinary leukotriene excretion in patients with cardiac ischemia. In vivo evidence for 5-lipoxygenase activation. Circulation 85, 230-236. Carter, G.W., Young, P.R., Albert, D.H., Bouska, J., Dyer, R., Bell, R.L., Summers, J.B., and Brooks, D.W. (1991). 5-lipoxygenase inhibitory activity of zileuton. The Journal of pharmacology and experimental therapeutics 256, 929-937. Chen, H., Qin, J., Wei, P., Zhang, J., Li, Q., Fu, L., Li, S., Ma, C., and Cong, B. (2009). Effects of leukotriene B4 and prostaglandin E2 on the differentiation of murine Foxp3+ T regulatory cells and Th17 cells. Prostaglandins, leukotrienes, and essential fatty acids 80, 195-200. Chen, L., Grabowski, K.A., Xin, J.P., Coleman, J., Huang, Z., Espiritu, B., Alkan, S., Xie, H.B., Zhu, Y., White, F.A., et al. (2004a). IL-4 induces differentiation and expansion of Th2 cytokineproducing eosinophils. Journal of immunology 172, 2059-2066. Chen, X., Li, N., Wang, S., Wu, N., Hong, J., Jiao, X., Krasna, M.J., Beer, D.G., and Yang, C.S. (2003). Leukotriene A4 hydrolase in rat and human esophageal adenocarcinomas and inhibitory effects of bestatin. Journal of the National Cancer Institute 95, 1053-1061. Chen, X., Wang, S., Wu, N., Sood, S., Wang, P., Jin, Z., Beer, D.G., Giordano, T.J., Lin, Y., Shih, W.C., et al. (2004b). Overexpression of 5-lipoxygenase in rat and human esophageal adenocarcinoma and inhibitory effects of zileuton and celecoxib on carcinogenesis. Clinical cancer research : an official journal of the American Association for Cancer Research 10, 67036709. Chen, Z., Siu, B., Ho, Y.S., Vincent, R., Chua, C.C., Hamdy, R.C., and Chua, B.H. (1998). Overexpression of MnSOD protects against myocardial ischemia/reperfusion injury in transgenic mice. Journal of molecular and cellular cardiology 30, 2281-2289. Chibana, K., Ishii, Y., Asakura, T., and Fukuda, T. (2003). Up-regulation of cysteinyl leukotriene 1 receptor by IL-13 enables human lung fibroblasts to respond to leukotriene C4 and produce eotaxin. Journal of immunology 170, 4290-4295. Chong, A.J., Shimamoto, A., Hampton, C.R., Takayama, H., Spring, D.J., Rothnie, C.L., Yada, M., Pohlman, T.H., and Verrier, E.D. (2004). Toll-like receptor 4 mediates ischemia/reperfusion injury of the heart. The Journal of thoracic and cardiovascular surgery 128, 170-179. 160 Christie, P.E., Smith, C.M., and Lee, T.H. (1991). The potent and selective sulfidopeptide leukotriene antagonist, SK&F 104353, inhibits aspirin-induced asthma. The American review of respiratory disease 144, 957-958. Christmas, P., Weber, B.M., McKee, M., Brown, D., and Soberman, R.J. (2002). Membrane localization and topology of leukotriene C4 synthase. The Journal of biological chemistry 277, 28902-28908. Chung, J.W., Kim, G.Y., Mun, Y.C., Ahn, J.Y., Seong, C.M., and Kim, J.H. (2005). Leukotriene B4 pathway regulates the fate of the hematopoietic stem cells. Experimental & molecular medicine 37, 45-50. Ciana, P., Fumagalli, M., Trincavelli, M.L., Verderio, C., Rosa, P., Lecca, D., Ferrario, S., Parravicini, C., Capra, V., Gelosa, P., et al. (2006). The orphan receptor GPR17 identified as a new dual uracil nucleotides/cysteinyl-leukotrienes receptor. The EMBO journal 25, 4615-4627. Clark, J.D., Schievella, A.R., Nalefski, E.A., and Lin, L.L. (1995). Cytosolic phospholipase A2. Journal of lipid mediators and cell signalling 12, 83-117. Coutts, I.G.C., Edwards, M., and Richards, D.J. (1981). Mild selective hydrolysis of the methyl esters of some ortho-substituted aromatic carboxylic acids. Synthesis 1981, 487–489. Covin, R.B., Brock, T.G., Bailie, M.B., and Peters-Golden, M. (1998). Altered expression and localization of 5-lipoxygenase accompany macrophage differentiation in the lung. The American journal of physiology 275, L303-310. Cowburn, A.S., Holgate, S.T., and Sampson, A.P. (1999). IL-5 increases expression of 5lipoxygenase-activating protein and translocates 5-lipoxygenase to the nucleus in human blood eosinophils. Journal of immunology 163, 456-465. Dahinden, C.A., Clancy, R.M., Gross, M., Chiller, J.M., and Hugli, T.E. (1985). Leukotriene C4 production by murine mast cells: evidence of a role for extracellular leukotriene A4. Proceedings of the National Academy of Sciences of the United States of America 82, 6632-6636. Dahlen, S.E. (2006). Treatment of asthma with antileukotrienes: first line or last resort therapy? European journal of pharmacology 533, 40-56. Dahlen, S.E., Bjork, J., Hedqvist, P., Arfors, K.E., Hammarstrom, S., Lindgren, J.A., and Samuelsson, B. (1981). Leukotrienes promote plasma leakage and leukocyte adhesion in postcapillary venules: in vivo effects with relevance to the acute inflammatory response. Proceedings of the National Academy of Sciences of the United States of America 78, 38873891. Dahlen, S.E., Hedqvist, P., Hammarstrom, S., and Samuelsson, B. (1980). Leukotrienes are potent constrictors of human bronchi. Nature 288, 484-486. 161 Daniele, S., Lecca, D., Trincavelli, M.L., Ciampi, O., Abbracchio, M.P., and Martini, C. (2010). Regulation of PC12 cell survival and differentiation by the new P2Y-like receptor GPR17. Cellular signalling 22, 697-706. Daniele, S., Trincavelli, M.L., Gabelloni, P., Lecca, D., Rosa, P., Abbracchio, M.P., and Martini, C. (2011). Agonist-induced desensitization/resensitization of human G protein-coupled receptor 17: a functional cross-talk between purinergic and cysteinyl-leukotriene ligands. The Journal of pharmacology and experimental therapeutics 338, 559-567. Datta, Y.H., Romano, M., Jacobson, B.C., Golan, D.E., Serhan, C.N., and Ewenstein, B.M. (1995). Peptido-leukotrienes are potent agonists of von Willebrand factor secretion and Pselectin surface expression in human umbilical vein endothelial cells. Circulation 92, 3304-3311. Davidson, A.B., Lee, T.H., Scanlon, P.D., Solway, J., McFadden, E.R., Jr., Ingram, R.H., Jr., Corey, E.J., Austen, K.F., and Drazen, J.M. (1987). Bronchoconstrictor effects of leukotriene E4 in normal and asthmatic subjects. The American review of respiratory disease 135, 333-337. Davies, M.J., Woolf, N., and Robertson, W.B. (1976). Pathology of acute myocardial infarction with particular reference to occlusive coronary thrombi. British heart journal 38, 659-664. De Palma, M., Venneri, M.A., Galli, R., Sergi Sergi, L., Politi, L.S., Sampaolesi, M., and Naldini, L. (2005). Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer cell 8, 211-226. Demitsu, T., Katayama, H., Saito-Taki, T., Yaoita, H., and Nakano, M. (1989). Phagocytosis and bactericidal action of mouse peritoneal macrophages treated with leukotriene B4. International journal of immunopharmacology 11, 801-808. Di Gennaro, A., Carnini, C., Buccellati, C., Ballerio, R., Zarini, S., Fumagalli, F., Viappiani, S., Librizzi, L., Hernandez, A., Murphy, R.C., et al. (2004). Cysteinyl-leukotrienes receptor activation in brain inflammatory reactions and cerebral edema formation: a role for transcellular biosynthesis of cysteinyl-leukotrienes. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 18, 842-844. Di Gennaro, A., Kenne, E., Wan, M., Soehnlein, O., Lindbom, L., and Haeggstrom, J.Z. (2009). Leukotriene B4-induced changes in vascular permeability are mediated by neutrophil release of heparin-binding protein (HBP/CAP37/azurocidin). FASEB journal : official publication of the Federation of American Societies for Experimental Biology 23, 1750-1757. Di Gennaro, A., Wagsater, D., Mayranpaa, M.I., Gabrielsen, A., Swedenborg, J., Hamsten, A., Samuelsson, B., Eriksson, P., and Haeggstrom, J.Z. (2010). Increased expression of leukotriene C4 synthase and predominant formation of cysteinyl-leukotrienes in human abdominal aortic aneurysm. Proceedings of the National Academy of Sciences of the United States of America 107, 21093-21097. Di Lisa, F., and Bernardi, P. (2006). Mitochondria and ischemia-reperfusion injury of the heart: fixing a hole. Cardiovascular research 70, 191-199. 162 Diamant, Z., Grootendorst, D.C., Veselic-Charvat, M., Timmers, M.C., De Smet, M., Leff, J.A., Seidenberg, B.C., Zwinderman, A.H., Peszek, I., and Sterk, P.J. (1999). The effect of montelukast (MK-0476), a cysteinyl leukotriene receptor antagonist, on allergen-induced airway responses and sputum cell counts in asthma. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 29, 42-51. Ding, Q., Fang, S.H., Zhou, Y., Zhang, L.H., Zhang, W.P., Chen, Z., and Wei, E.Q. (2007). Cysteinyl leukotriene receptor 1 partially mediates brain cryoinjury in mice. Acta pharmacologica Sinica 28, 945-952. Ding, X.Z., Talamonti, M.S., Bell, R.H., Jr., and Adrian, T.E. (2005). A novel anti-pancreatic cancer agent, LY293111. Anti-cancer drugs 16, 467-473. Dixon, R.A., Diehl, R.E., Opas, E., Rands, E., Vickers, P.J., Evans, J.F., Gillard, J.W., and Miller, D.K. (1990). Requirement of a 5-lipoxygenase-activating protein for leukotriene synthesis. Nature 343, 282-284. Dockhorn, R.J., Baumgartner, R.A., Leff, J.A., Noonan, M., Vandormael, K., Stricker, W., Weinland, D.E., and Reiss, T.F. (2000). Comparison of the effects of intravenous and oral montelukast on airway function: a double blind, placebo controlled, three period, crossover study in asthmatic patients. Thorax 55, 260-265. Drazen, J.M., Venugopalan, C.S., Austen, K.F., Brion, F., and Corey, E.J. (1982). Effects of leukotriene E on pulmonary mechanics in the guinea pig. The American review of respiratory disease 125, 290-294. Dwyer, J.H., Allayee, H., Dwyer, K.M., Fan, J., Wu, H., Mar, R., Lusis, A.J., and Mehrabian, M. (2004). Arachidonate 5-lipoxygenase promoter genotype, dietary arachidonic acid, and atherosclerosis. The New England journal of medicine 350, 29-37. Early, S.B., Barekzi, E., Negri, J., Hise, K., Borish, L., and Steinke, J.W. (2007). Concordant modulation of cysteinyl leukotriene receptor expression by IL-4 and IFN-gamma on peripheral immune cells. American journal of respiratory cell and molecular biology 36, 715-720. Eaton, A., Nagy, E., Pacault, M., Fauconnier, J., and Back, M. (2012). Cysteinyl leukotriene signaling through perinuclear CysLT(1) receptors on vascular smooth muscle cells transduces nuclear calcium signaling and alterations of gene expression. Journal of molecular medicine 90, 1223-1231. Edenius, C., Heidvall, K., and Lindgren, J.A. (1988). Novel transcellular interaction: conversion of granulocyte-derived leukotriene A4 to cysteinyl-containing leukotrienes by human platelets. European journal of biochemistry / FEBS 178, 81-86. Eguchi, M., Kim, Y.H., Kang, K.W., Shim, C.Y., Jang, Y., Dorval, T., Kim, K.J., and Sweeney, G. (2012). Ischemia-reperfusion injury leads to distinct temporal cardiac remodeling in normal versus diabetic mice. PloS one 7, e30450. 163 Eming, S.A., Krieg, T., and Davidson, J.M. (2007). Inflammation in wound repair: molecular and cellular mechanisms. The Journal of investigative dermatology 127, 514-525. Entman, M.L., Youker, K., Shoji, T., Kukielka, G., Shappell, S.B., Taylor, A.A., and Smith, C.W. (1992). Neutrophil induced oxidative injury of cardiac myocytes. A compartmented system requiring CD11b/CD18-ICAM-1 adherence. The Journal of clinical investigation 90, 1335-1345. Espinosa, K., Bosse, Y., Stankova, J., and Rola-Pleszczynski, M. (2003). CysLT1 receptor upregulation by TGF-beta and IL-13 is associated with bronchial smooth muscle cell proliferation in response to LTD4. The Journal of allergy and clinical immunology 111, 10321040. Evans, J.F. (2003). The cysteinyl leukotriene receptors. Prostaglandins, leukotrienes, and essential fatty acids 69, 117-122. Fabre, J.E., Goulet, J.L., Riche, E., Nguyen, M., Coggins, K., Offenbacher, S., and Koller, B.H. (2002). Transcellular biosynthesis contributes to the production of leukotrienes during inflammatory responses in vivo. The Journal of clinical investigation 109, 1373-1380. Faith, A., Fernandez, M.H., Caulfield, J., Loke, T.K., Corrigan, C., O'Connor, B., Lee, T.H., and Hawrylowicz, C.M. (2008). Role of cysteinyl leukotrienes in human allergen-specific Th2 responses induced by granulocyte macrophage-colony stimulating factor. Allergy 63, 168-175. Fang, S.H., Wei, E.Q., Zhou, Y., Wang, M.L., Zhang, W.P., Yu, G.L., Chu, L.S., and Chen, Z. (2006). Increased expression of cysteinyl leukotriene receptor-1 in the brain mediates neuronal damage and astrogliosis after focal cerebral ischemia in rats. Neuroscience 140, 969-979. Fang, S.H., Zhou, Y., Chu, L.S., Zhang, W.P., Wang, M.L., Yu, G.L., Peng, F., and Wei, E.Q. (2007). Spatio-temporal expression of cysteinyl leukotriene receptor-2 mRNA in rat brain after focal cerebral ischemia. Neuroscience letters 412, 78-83. Fegn, L., and Wang, Z. (2009). Topical chemoprevention of skin cancer in mice, using combined inhibitors of 5-lipoxygenase and cyclo-oxygenase-2. The Journal of laryngology and otology 123, 880-884. Feinmark, S.J., and Cannon, P.J. (1986). Endothelial cell leukotriene C4 synthesis results from intercellular transfer of leukotriene A4 synthesized by polymorphonuclear leukocytes. The Journal of biological chemistry 261, 16466-16472. Feldberg, W., Holden, H.F., and Kellaway, C.H. (1938). The formation of lysocithin and of a muscle-stimulating substance by snake venoms. The Journal of physiology 94, 232-248. Figueroa, D.J., Borish, L., Baramki, D., Philip, G., Austin, C.P., and Evans, J.F. (2003). Expression of cysteinyl leukotriene synthetic and signalling proteins in inflammatory cells in active seasonal allergic rhinitis. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 33, 1380-1388. 164 Figueroa, D.J., Breyer, R.M., Defoe, S.K., Kargman, S., Daugherty, B.L., Waldburger, K., Liu, Q., Clements, M., Zeng, Z., O'Neill, G.P., et al. (2001). Expression of the cysteinyl leukotriene 1 receptor in normal human lung and peripheral blood leukocytes. American journal of respiratory and critical care medicine 163, 226-233. Fischer, A.R., Rosenberg, M.A., Lilly, C.M., Callery, J.C., Rubin, P., Cohn, J., White, M.V., Igarashi, Y., Kaliner, M.A., Drazen, J.M., et al. (1994). Direct evidence for a role of the mast cell in the nasal response to aspirin in aspirin-sensitive asthma. The Journal of allergy and clinical immunology 94, 1046-1056. Fitzpatrick, F., Liggett, W., McGee, J., Bunting, S., Morton, D., and Samuelsson, B. (1984). Metabolism of leukotriene A4 by human erythrocytes. A novel cellular source of leukotriene B4. The Journal of biological chemistry 259, 11403-11407. Flamand, L., Tremblay, M.J., and Borgeat, P. (2007a). Leukotriene B4 triggers the in vitro and in vivo release of potent antimicrobial agents. Journal of immunology 178, 8036-8045. Flamand, N., Lefebvre, J., Surette, M.E., Picard, S., and Borgeat, P. (2006). Arachidonic acid regulates the translocation of 5-lipoxygenase to the nuclear membranes in human neutrophils. The Journal of biological chemistry 281, 129-136. Flamand, N., Mancuso, P., Serezani, C.H., and Brock, T.G. (2007b). Leukotrienes: mediators that have been typecast as villains. Cellular and molecular life sciences : CMLS 64, 2657-2670. Folco, G., and Murphy, R.C. (2006). Eicosanoid transcellular biosynthesis: from cell-cell interactions to in vivo tissue responses. Pharmacological reviews 58, 375-388. Ford-Hutchinson, A.W., Bray, M.A., Doig, M.V., Shipley, M.E., and Smith, M.J. (1980). Leukotriene B, a potent chemokinetic and aggregating substance released from polymorphonuclear leukocytes. Nature 286, 264-265. Foster, H.R., Fuerst, E., Lee, T.H., Cousins, D.J., and Woszczek, G. (2013). Characterisation of P2Y(12) receptor responsiveness to cysteinyl leukotrienes. PloS one 8, e58305. Fradin, A., Zirrolli, J.A., Maclouf, J., Vausbinder, L., Henson, P.M., and Murphy, R.C. (1989). Platelet-activating factor and leukotriene biosynthesis in whole blood. A model for the study of transcellular arachidonate metabolism. Journal of immunology 143, 3680-3685. Frangogiannis, N.G. (2012). Regulation of the inflammatory response in cardiac repair. Circulation research 110, 159-173. Frangogiannis, N.G., Smith, C.W., and Entman, M.L. (2002). The inflammatory response in myocardial infarction. Cardiovascular research 53, 31-47. Frantz, S., Hofmann, U., Fraccarollo, D., Schafer, A., Kranepuhl, S., Hagedorn, I., Nieswandt, B., Nahrendorf, M., Wagner, H., Bayer, B., et al. (2013). Monocytes/macrophages prevent healing defects and left ventricular thrombus formation after myocardial infarction. FASEB 165 journal : official publication of the Federation of American Societies for Experimental Biology 27, 871-881. Fregonese, L., Silvestri, M., Sabatini, F., and Rossi, G.A. (2002). Cysteinyl leukotrienes induce human eosinophil locomotion and adhesion molecule expression via a CysLT1 receptormediated mechanism. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 32, 745-750. Freiberg, J.J., Dahl, M., Tybjaerg-Hansen, A., Grande, P., and Nordestgaard, B.G. (2009). Leukotriene C4 synthase and ischemic cardiovascular disease and obstructive pulmonary disease in 13,000 individuals. Journal of molecular and cellular cardiology 46, 579-586. Frey, N., Katus, H.A., Olson, E.N., and Hill, J.A. (2004). Hypertrophy of the heart: a new therapeutic target? Circulation 109, 1580-1589. Friedrich, E.B., Tager, A.M., Liu, E., Pettersson, A., Owman, C., Munn, L., Luster, A.D., and Gerszten, R.E. (2003). Mechanisms of leukotriene B4--triggered monocyte adhesion. Arteriosclerosis, thrombosis, and vascular biology 23, 1761-1767. Frieri, M., Therattil, J., Wang, S.F., Huang, C.Y., and Wang, Y.C. (2003). Montelukast inhibits interleukin-5 mRNA expression and cysteinyl leukotriene production in ragweed and mitestimulated peripheral blood mononuclear cells from patients with asthma. Allergy and asthma proceedings : the official journal of regional and state allergy societies 24, 359-366. Funk, C.D. (2001). Prostaglandins and leukotrienes: advances in eicosanoid biology. Science 294, 1871-1875. Funk, C.D. (2005). Leukotriene modifiers as potential therapeutics for cardiovascular disease. Nature reviews Drug discovery 4, 664-672. Funk, C.D., Chen, X.S., Kurre, U., and Griffis, G. (1995). Leukotriene-deficient mice generated by targeted disruption of the 5-lipoxygenase gene. Advances in prostaglandin, thromboxane, and leukotriene research 23, 145-150. Gaddy, J.N., Margolskee, D.J., Bush, R.K., Williams, V.C., and Busse, W.W. (1992). Bronchodilation with a potent and selective leukotriene D4 (LTD4) receptor antagonist (MK571) in patients with asthma. The American review of respiratory disease 146, 358-363. Gauvreau, G.M., Parameswaran, K.N., Watson, R.M., and O'Byrne, P.M. (2001). Inhaled leukotriene E(4), but not leukotriene D(4), increased airway inflammatory cells in subjects with atopic asthma. American journal of respiratory and critical care medicine 164, 1495-1500. Ge, S., Zhou, G., Cheng, S., Liu, D., Xu, J., Xu, G., and Liu, X. (2009). Anti-atherogenic effects of montelukast associated with reduced MCP-1 expression in a rabbit carotid balloon injury model. Atherosclerosis 205, 74-79. 166 Gimbrone, M.A., Jr., Brock, A.F., and Schafer, A.I. (1984). Leukotriene B4 stimulates polymorphonuclear leukocyte adhesion to cultured vascular endothelial cells. The Journal of clinical investigation 74, 1552-1555. Glover, S., de Carvalho, M.S., Bayburt, T., Jonas, M., Chi, E., Leslie, C.C., and Gelb, M.H. (1995). Translocation of the 85-kDa phospholipase A2 from cytosol to the nuclear envelope in rat basophilic leukemia cells stimulated with calcium ionophore or IgE/antigen. The Journal of biological chemistry 270, 15359-15367. Gonsalves, C.S., and Kalra, V.K. (2010). Hypoxia-mediated expression of 5-lipoxygenaseactivating protein involves HIF-1alpha and NF-kappaB and microRNAs 135a and 199a-5p. Journal of immunology 184, 3878-3888. Goodarzi, K., Goodarzi, M., Tager, A.M., Luster, A.D., and von Andrian, U.H. (2003). Leukotriene B4 and BLT1 control cytotoxic effector T cell recruitment to inflamed tissues. Nature immunology 4, 965-973. Graham, I., Atar, D., Borch-Johnsen, K., Boysen, G., Burell, G., Cifkova, R., Dallongeville, J., De Backer, G., Ebrahim, S., Gjelsvik, B., et al. (2007). European guidelines on cardiovascular disease prevention in clinical practice: executive summary: Fourth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (Constituted by representatives of nine societies and by invited experts). European heart journal 28, 2375-2414. Grimminger, F., Kreusler, B., Schneider, U., Becker, G., and Seeger, W. (1990). Influence of microvascular adherence on neutrophil leukotriene generation. Evidence for cooperative eicosanoid synthesis. Journal of immunology 144, 1866-1872. Grimminger, F., Menger, M., Becker, G., and Seeger, W. (1988). Potentiation of leukotriene production following sequestration of neutrophils in isolated lungs: indirect evidence for intercellular leukotriene A4 transfer. Blood 72, 1687-1692. Grimminger, F., Sibelius, U., and Seeger, W. (1991). Amplification of LTB4 generation in AMPMN cocultures: transcellular 5-lipoxygenase metabolism. The American journal of physiology 261, L195-203. Gronert, K., Martinsson-Niskanen, T., Ravasi, S., Chiang, N., and Serhan, C.N. (2001). Selectivity of recombinant human leukotriene D(4), leukotriene B(4), and lipoxin A(4) receptors with aspirin-triggered 15-epi-LXA(4) and regulation of vascular and inflammatory responses. The American journal of pathology 158, 3-9. Gunning, W.T., Kramer, P.M., Steele, V.E., and Pereira, M.A. (2002). Chemoprevention by lipoxygenase and leukotriene pathway inhibitors of vinyl carbamate-induced lung tumors in mice. Cancer research 62, 4199-4201. Haeggstrom, J.Z. (2004). Leukotriene A4 hydrolase/aminopeptidase, the gatekeeper of chemotactic leukotriene B4 biosynthesis. The Journal of biological chemistry 279, 50639-50642. 167 Haeggstrom, J.Z., and Funk, C.D. (2011). Lipoxygenase and leukotriene pathways: biochemistry, biology, and roles in disease. Chemical reviews 111, 5866-5898. Hafstrom, I., Palmblad, J., Malmsten, C.L., Radmark, O., and Samuelsson, B. (1981). Leukotriene B4--a stereospecific stimulator for release of lysosomal enzymes from neutrophils. FEBS letters 130, 146-148. Hahn, R.A., MacDonald, B.R., Morgan, E., Potts, B.D., Parli, C.J., Rinkema, L.E., Whitesitt, C.A., and Marshall, W.S. (1992). Evaluation of LY203647 on cardiovascular leukotriene D4 receptors and myocardial reperfusion injury. The Journal of pharmacology and experimental therapeutics 260, 979-989. Halestrap, A.P., Clarke, S.J., and Javadov, S.A. (2004). Mitochondrial permeability transition pore opening during myocardial reperfusion--a target for cardioprotection. Cardiovascular research 61, 372-385. Harada, K., Sugaya, T., Murakami, K., Yazaki, Y., and Komuro, I. (1999). Angiotensin II type 1A receptor knockout mice display less left ventricular remodeling and improved survival after myocardial infarction. Circulation 100, 2093-2099. Harter, M., Erguden, J., Wunder, F., Tinel, H., Kobberling, J., Becker, E.M., Munter, K., Schlemmer, K.H., Kast, R., Kolkhof, P., et al. (2009). Isophthalic acid derivatives (United States: Bayer Healthcare AG). Hasegawa, S., Ichiyama, T., Hashimoto, K., Suzuki, Y., Hirano, R., Fukano, R., and Furukawa, S. (2010). Functional expression of cysteinyl leukotriene receptors on human platelets. Platelets 21, 253-259. Hashimoto, H., Miyazawa, K., Hagiwara, M., Miyasaka, K., and Nakashima, M. (1990). Beneficial effects of a new 5-lipoxygenase inhibitor on occlusion- and occlusion-reperfusioninduced myocardial injury. Arzneimittel-Forschung 40, 126-129. Hashimoto, K., Ichiyama, T., Hasegawa, M., Hasegawa, S., Matsubara, T., and Furukawa, S. (2009). Cysteinyl leukotrienes induce monocyte chemoattractant protein-1 in human monocyte/macrophages via mitogen-activated protein kinase and nuclear factor-kappaB pathways. International archives of allergy and immunology 149, 275-282. He, W., Miao, F.J., Lin, D.C., Schwandner, R.T., Wang, Z., Gao, J., Chen, J.L., Tian, H., and Ling, L. (2004). Citric acid cycle intermediates as ligands for orphan G-protein-coupled receptors. Nature 429, 188-193. Hebert, M.J., Takano, T., Holthofer, H., and Brady, H.R. (1996). Sequential morphologic events during apoptosis of human neutrophils. Modulation by lipoxygenase-derived eicosanoids. Journal of immunology 157, 3105-3115. Heidenreich, P.A., Trogdon, J.G., Khavjou, O.A., Butler, J., Dracup, K., Ezekowitz, M.D., Finkelstein, E.A., Hong, Y., Johnston, S.C., Khera, A., et al. (2011). Forecasting the future of 168 cardiovascular disease in the United States: a policy statement from the American Heart Association. Circulation 123, 933-944. Heise, C.E., O'Dowd, B.F., Figueroa, D.J., Sawyer, N., Nguyen, T., Im, D.S., Stocco, R., Bellefeuille, J.N., Abramovitz, M., Cheng, R., et al. (2000). Characterization of the human cysteinyl leukotriene 2 receptor. The Journal of biological chemistry 275, 30531-30536. Helgadottir, A., Gretarsdottir, S., St Clair, D., Manolescu, A., Cheung, J., Thorleifsson, G., Pasdar, A., Grant, S.F., Whalley, L.J., Hakonarson, H., et al. (2005). Association between the gene encoding 5-lipoxygenase-activating protein and stroke replicated in a Scottish population. American journal of human genetics 76, 505-509. Helgadottir, A., Manolescu, A., Helgason, A., Thorleifsson, G., Thorsteinsdottir, U., Gudbjartsson, D.F., Gretarsdottir, S., Magnusson, K.P., Gudmundsson, G., Hicks, A., et al. (2006). A variant of the gene encoding leukotriene A4 hydrolase confers ethnicity-specific risk of myocardial infarction. Nature genetics 38, 68-74. Helgadottir, A., Manolescu, A., Thorleifsson, G., Gretarsdottir, S., Jonsdottir, H., Thorsteinsdottir, U., Samani, N.J., Gudmundsson, G., Grant, S.F., Thorgeirsson, G., et al. (2004). The gene encoding 5-lipoxygenase activating protein confers risk of myocardial infarction and stroke. Nature genetics 36, 233-239. Henderson, W.R., Jr., Tang, L.O., Chu, S.J., Tsao, S.M., Chiang, G.K., Jones, F., Jonas, M., Pae, C., Wang, H., and Chi, E.Y. (2002). A role for cysteinyl leukotrienes in airway remodeling in a mouse asthma model. American journal of respiratory and critical care medicine 165, 108-116. Hennig, R., Ventura, J., Segersvard, R., Ward, E., Ding, X.Z., Rao, S.M., Jovanovic, B.D., Iwamura, T., Talamonti, M.S., Bell, R.H., Jr., et al. (2005). LY293111 improves efficacy of gemcitabine therapy on pancreatic cancer in a fluorescent orthotopic model in athymic mice. Neoplasia 7, 417-425. Hernandez, L.A., Grisham, M.B., Twohig, B., Arfors, K.E., Harlan, J.M., and Granger, D.N. (1987). Role of neutrophils in ischemia-reperfusion-induced microvascular injury. The American journal of physiology 253, H699-703. Heymans, S., Luttun, A., Nuyens, D., Theilmeier, G., Creemers, E., Moons, L., Dyspersin, G.D., Cleutjens, J.P., Shipley, M., Angellilo, A., et al. (1999). Inhibition of plasminogen activators or matrix metalloproteinases prevents cardiac rupture but impairs therapeutic angiogenesis and causes cardiac failure. Nature medicine 5, 1135-1142. Hirabayashi, T., Murayama, T., and Shimizu, T. (2004). Regulatory mechanism and physiological role of cytosolic phospholipase A2. Biological & pharmaceutical bulletin 27, 1168-1173. Hock, C.E., Beck, L.D., and Papa, L.A. (1992). Peptide leukotriene receptor antagonism in myocardial ischaemia and reperfusion. Cardiovascular research 26, 1206-1211. 169 Hofmann, U., Beyersdorf, N., Weirather, J., Podolskaya, A., Bauersachs, J., Ertl, G., Kerkau, T., and Frantz, S. (2012). Activation of CD4+ T lymphocytes improves wound healing and survival after experimental myocardial infarction in mice. Circulation 125, 1652-1663. Horrillo, R., Gonzalez-Periz, A., Martinez-Clemente, M., Lopez-Parra, M., Ferre, N., Titos, E., Moran-Salvador, E., Deulofeu, R., Arroyo, V., and Claria, J. (2010). 5-lipoxygenase activating protein signals adipose tissue inflammation and lipid dysfunction in experimental obesity. Journal of immunology 184, 3978-3987. Hoshino, M., Izumi, T., and Shimizu, T. (1998). Leukotriene D4 activates mitogen-activated protein kinase through a protein kinase Calpha-Raf-1-dependent pathway in human monocytic leukemia THP-1 cells. The Journal of biological chemistry 273, 4878-4882. Huang, H., Zeng, Z., Li, J., Zhang, L., and Chen, Y. (2010). Variants of arachidonate 5lipoxygenase-activating protein (ALOX5AP) gene and risk of coronary heart disease: A metaanalysis. Archives of medical research 41, 634-641. Huang, L., Zhao, A., Wong, F., Ayala, J.M., Struthers, M., Ujjainwalla, F., Wright, S.D., Springer, M.S., Evans, J., and Cui, J. (2004). Leukotriene B4 strongly increases monocyte chemoattractant protein-1 in human monocytes. Arteriosclerosis, thrombosis, and vascular biology 24, 1783-1788. Huang, W.W., Garcia-Zepeda, E.A., Sauty, A., Oettgen, H.C., Rothenberg, M.E., and Luster, A.D. (1998). Molecular and biological characterization of the murine leukotriene B4 receptor expressed on eosinophils. The Journal of experimental medicine 188, 1063-1074. Huang, X.J., Zhang, W.P., Li, C.T., Shi, W.Z., Fang, S.H., Lu, Y.B., Chen, Z., and Wei, E.Q. (2008). Activation of CysLT receptors induces astrocyte proliferation and death after oxygenglucose deprivation. Glia 56, 27-37. Hui, Y., Cheng, Y., Smalera, I., Jian, W., Goldhahn, L., Fitzgerald, G.A., and Funk, C.D. (2004). Directed vascular expression of human cysteinyl leukotriene 2 receptor modulates endothelial permeability and systemic blood pressure. Circulation 110, 3360-3366. Hui, Y., Yang, G., Galczenski, H., Figueroa, D.J., Austin, C.P., Copeland, N.G., Gilbert, D.J., Jenkins, N.A., and Funk, C.D. (2001). The murine cysteinyl leukotriene 2 (CysLT2) receptor. cDNA and genomic cloning, alternative splicing, and in vitro characterization. The Journal of biological chemistry 276, 47489-47495. Ichiyama, T., Hasegawa, M., Hashimoto, K., Matsushige, T., Hirano, R., and Furukawa, S. (2009). Cysteinyl leukotrienes induce macrophage inflammatory protein-1 in human monocytes/macrophages. International archives of allergy and immunology 148, 147-153. Ichiyama, T., Hasegawa, M., Ueno, Y., Makata, H., Matsubara, T., and Furukawa, S. (2005). Cysteinyl leukotrienes induce monocyte chemoattractant protein 1 in human monocytes/macrophages. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 35, 1214-1219. 170 Ihara, A., Wada, K., Yoneda, M., Fujisawa, N., Takahashi, H., and Nakajima, A. (2007). Blockade of leukotriene B4 signaling pathway induces apoptosis and suppresses cell proliferation in colon cancer. Journal of pharmacological sciences 103, 24-32. Islam, S.A., Thomas, S.Y., Hess, C., Medoff, B.D., Means, T.K., Brander, C., Lilly, C.M., Tager, A.M., and Luster, A.D. (2006). The leukotriene B4 lipid chemoattractant receptor BLT1 defines antigen-primed T cells in humans. Blood 107, 444-453. Israel, E., Cohn, J., Dube, L., and Drazen, J.M. (1996). Effect of treatment with zileuton, a 5lipoxygenase inhibitor, in patients with asthma. A randomized controlled trial. Zileuton Clinical Trial Group. JAMA : the journal of the American Medical Association 275, 931-936. Ito, T., Toki, Y., Hieda, N., Okumura, K., Hashimoto, H., Ogawa, K., and Satake, T. (1989). Protective effects of a thromboxane synthetase inhibitor, a thromboxane antagonist, a lipoxygenase inhibitor and a leukotriene C4, D4 antagonist on myocardial injury caused by acute myocardial infarction in the canine heart. Japanese circulation journal 53, 1115-1121. Iversen, L., Kristensen, P., Gron, B., Ziboh, V.A., and Kragballe, K. (1994). Human epidermis transforms exogenous leukotriene A4 into peptide leukotrienes: possible role in transcellular metabolism. Archives of dermatological research 286, 261-266. Jawien, J., Gajda, M., Wolkow, P., Zuranska, J., Olszanecki, R., and Korbut, R. (2008). The effect of montelukast on atherogenesis in apoE/LDLR-double knockout mice. Journal of physiology and pharmacology : an official journal of the Polish Physiological Society 59, 633639. Jeong, C.H., Bode, A.M., Pugliese, A., Cho, Y.Y., Kim, H.G., Shim, J.H., Jeon, Y.J., Li, H., Jiang, H., and Dong, Z. (2009). [6]-Gingerol suppresses colon cancer growth by targeting leukotriene A4 hydrolase. Cancer research 69, 5584-5591. Jiang, W., Hall, S.R., Moos, M.P., Cao, R.Y., Ishii, S., Ogunyankin, K.O., Melo, L.G., and Funk, C.D. (2008). Endothelial cysteinyl leukotriene 2 receptor expression mediates myocardial ischemia-reperfusion injury. The American journal of pathology 172, 592-602. Jiang, Y., Borrelli, L.A., Kanaoka, Y., Bacskai, B.J., and Boyce, J.A. (2007). CysLT2 receptors interact with CysLT1 receptors and down-modulate cysteinyl leukotriene dependent mitogenic responses of mast cells. Blood 110, 3263-3270. Jiang, Y., Kanaoka, Y., Feng, C., Nocka, K., Rao, S., and Boyce, J.A. (2006). Cutting edge: Interleukin 4-dependent mast cell proliferation requires autocrine/intracrine cysteinyl leukotriene-induced signaling. Journal of immunology 177, 2755-2759. Jolly, S.R., Kane, W.J., Bailie, M.B., Abrams, G.D., and Lucchesi, B.R. (1984). Canine myocardial reperfusion injury. Its reduction by the combined administration of superoxide dismutase and catalase. Circulation research 54, 277-285. Jones, T.R., Labelle, M., Belley, M., Champion, E., Charette, L., Evans, J., Ford-Hutchinson, A.W., Gauthier, J.Y., Lord, A., Masson, P., et al. (1995). Pharmacology of montelukast sodium 171 (Singulair), a potent and selective leukotriene D4 receptor antagonist. Canadian journal of physiology and pharmacology 73, 191-201. Jung, K., Kim, P., Leuschner, F., Gorbatov, R., Kim, J.K., Ueno, T., Nahrendorf, M., and Yun, S.H. (2013). Endoscopic time-lapse imaging of immune cells in infarcted mouse hearts. Circulation research 112, 891-899. Jung, U., and Ley, K. (1999). Mice lacking two or all three selectins demonstrate overlapping and distinct functions for each selectin. Journal of immunology 162, 6755-6762. Jung, U., Norman, K.E., Scharffetter-Kochanek, K., Beaudet, A.L., and Ley, K. (1998). Transit time of leukocytes rolling through venules controls cytokine-induced inflammatory cell recruitment in vivo. The Journal of clinical investigation 102, 1526-1533. Kaetsu, Y., Yamamoto, Y., Sugihara, S., Matsuura, T., Igawa, G., Matsubara, K., Igawa, O., Shigemasa, C., and Hisatome, I. (2007). Role of cysteinyl leukotrienes in the proliferation and the migration of murine vascular smooth muscle cells in vivo and in vitro. Cardiovascular research 76, 160-166. Kamohara, M., Takasaki, J., Matsumoto, M., Matsumoto, S., Saito, T., Soga, T., Matsushime, H., and Furuichi, K. (2001). Functional characterization of cysteinyl leukotriene CysLT(2) receptor on human coronary artery smooth muscle cells. Biochemical and biophysical research communications 287, 1088-1092. Kanaoka, Y., Maekawa, A., and Austen, K.F. (2013). Identification of GPR99 Protein as a Potential Third Cysteinyl Leukotriene Receptor with a Preference for Leukotriene E4 Ligand. The Journal of biological chemistry 288, 10967-10972. Kanaoka, Y., Maekawa, A., Penrose, J.F., Austen, K.F., and Lam, B.K. (2001). Attenuated zymosan-induced peritoneal vascular permeability and IgE-dependent passive cutaneous anaphylaxis in mice lacking leukotriene C4 synthase. The Journal of biological chemistry 276, 22608-22613. Kato, Y., Fujisawa, T., Nishimori, H., Katsumata, H., Atsuta, J., Iguchi, K., and Kamiya, H. (2005). Leukotriene D4 induces production of transforming growth factor-beta1 by eosinophils. International archives of allergy and immunology 137 Suppl 1, 17-20. Katsuki, H., and Okuda, S. (1995). Arachidonic acid as a neurotoxic and neurotrophic substance. Progress in neurobiology 46, 607-636. Kellaway, C.H., and Trethewie, E.R. (1940). The liberation of a slow-reacting smooth musclestimulating substance in anaphylaxis. Experimental Physiology 30, 121-145. Kempf, T., Zarbock, A., Vestweber, D., and Wollert, K.C. (2012). Anti-inflammatory mechanisms and therapeutic opportunities in myocardial infarct healing. Journal of molecular medicine 90, 361-369. 172 Kim, G.Y., Lee, J.W., Cho, S.H., Seo, J.M., and Kim, J.H. (2009). Role of the low-affinity leukotriene B4 receptor BLT2 in VEGF-induced angiogenesis. Arteriosclerosis, thrombosis, and vascular biology 29, 915-920. Kim, M.H., Lee, Y.J., Kim, M.O., Kim, J.S., and Han, H.J. (2010). Effect of leukotriene D4 on mouse embryonic stem cell migration and proliferation: involvement of PI3K/Akt as well as GSK-3beta/beta-catenin signaling pathways. Journal of cellular biochemistry 111, 686-698. Kiwamoto, T., Ishii, Y., Morishima, Y., Yoh, K., Kikuchi, N., Haraguchi, N., Masuko, H., Kawaguchi, M., Nomura, A., Sakamoto, T., et al. (2011). Blockade of cysteinyl leukotriene-1 receptors suppresses airway remodelling in mice overexpressing GATA-3. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 41, 116-128. Kjeldsen, L., Bjerrum, O.W., Askaa, J., and Borregaard, N. (1992). Subcellular localization and release of human neutrophil gelatinase, confirming the existence of separate gelatinasecontaining granules. The Biochemical journal 287 ( Pt 2), 603-610. Kleikers, P.W., Wingler, K., Hermans, J.J., Diebold, I., Altenhofer, S., Radermacher, K.A., Janssen, B., Gorlach, A., and Schmidt, H.H. (2012). NADPH oxidases as a source of oxidative stress and molecular target in ischemia/reperfusion injury. Journal of molecular medicine 90, 1391-1406. Kloner, R.A., and Jennings, R.B. (2001). Consequences of brief ischemia: stunning, preconditioning, and their clinical implications: part 1. Circulation 104, 2981-2989. Kolaczkowska, E., and Kubes, P. (2013). Neutrophil recruitment and function in health and inflammation. Nature reviews Immunology 13, 159-175. Kostulas, K., Gretarsdottir, S., Kostulas, V., Manolescu, A., Helgadottir, A., Thorleifsson, G., Gudmundsson, L.J., Thorsteinsdottir, U., Gulcher, J.R., Stefansson, K., et al. (2007). PDE4D and ALOX5AP genetic variants and risk for Ischemic Cerebrovascular Disease in Sweden. Journal of the neurological sciences 263, 113-117. Krell, R.D., Aharony, D., Buckner, C.K., Keith, R.A., Kusner, E.J., Snyder, D.W., Bernstein, P.R., Matassa, V.G., Yee, Y.K., Brown, F.J., et al. (1990). The preclinical pharmacology of ICI 204,219. A peptide leukotriene antagonist. The American review of respiratory disease 141, 978987. Kruh, G.D., and Belinsky, M.G. (2003). The MRP family of drug efflux pumps. Oncogene 22, 7537-7552. Kubes, P., Suzuki, M., and Granger, D.N. (1991). Nitric oxide: an endogenous modulator of leukocyte adhesion. Proceedings of the National Academy of Sciences of the United States of America 88, 4651-4655. Kumar, A.G., Ballantyne, C.M., Michael, L.H., Kukielka, G.L., Youker, K.A., Lindsey, M.L., Hawkins, H.K., Birdsall, H.H., MacKay, C.R., LaRosa, G.J., et al. (1997). Induction of 173 monocyte chemoattractant protein-1 in the small veins of the ischemic and reperfused canine myocardium. Circulation 95, 693-700. Kunkel, E.J., and Ley, K. (1996). Distinct phenotype of E-selectin-deficient mice. E-selectin is required for slow leukocyte rolling in vivo. Circulation research 79, 1196-1204. Ladilov, Y.V., Siegmund, B., and Piper, H.M. (1995). Protection of reoxygenated cardiomyocytes against hypercontracture by inhibition of Na+/H+ exchange. The American journal of physiology 268, H1531-1539. Lam, B.K., Penrose, J.F., Xu, K., and Austen, K.F. (1995). Leukotriene C4 synthase. Journal of lipid mediators and cell signalling 12, 333-341. Lamoureux, J., Stankova, J., and Rola-Pleszczynski, M. (2006). Leukotriene D4 enhances immunoglobulin production in CD40-activated human B lymphocytes. The Journal of allergy and clinical immunology 117, 924-930. Larre, S., Tran, N., Fan, C., Hamadeh, H., Champigneulles, J., Azzouzi, R., Cussenot, O., Mangin, P., and Olivier, J.L. (2008). PGE2 and LTB4 tissue levels in benign and cancerous prostates. Prostaglandins & other lipid mediators 87, 14-19. Lee, E., Robertson, T., Smith, J., and Kilfeather, S. (2000). Leukotriene receptor antagonists and synthesis inhibitors reverse survival in eosinophils of asthmatic individuals. American journal of respiratory and critical care medicine 161, 1881-1886. Lee, J.D., Lee, T.H., Huang, Y.C., Chang, Y.J., Chang, C.H., Hsu, H.L., Lin, Y.H., Wu, C.Y., Lee, M., Huang, Y.C., et al. (2011). ALOX5AP genetic variants and risk of atherothrombotic stroke in the Taiwanese population. Journal of clinical neuroscience : official journal of the Neurosurgical Society of Australasia 18, 1634-1638. Lee, K.S., Kim, S.R., Park, H.S., Park, S.J., Min, K.H., Lee, K.Y., Jin, S.M., and Lee, Y.C. (2007). Cysteinyl leukotriene upregulates IL-11 expression in allergic airway disease of mice. The Journal of allergy and clinical immunology 119, 141-149. Lee, T.H., Austen, K.F., Corey, E.J., and Drazen, J.M. (1984). Leukotriene E4-induced airway hyperresponsiveness of guinea pig tracheal smooth muscle to histamine and evidence for three separate sulfidopeptide leukotriene receptors. Proceedings of the National Academy of Sciences of the United States of America 81, 4922-4925. Leier, I., Jedlitschky, G., Buchholz, U., Cole, S.P., Deeley, R.G., and Keppler, D. (1994). The MRP gene encodes an ATP-dependent export pump for leukotriene C4 and structurally related conjugates. The Journal of biological chemistry 269, 27807-27810. Leikauf, G.D., Claesson, H.E., Doupnik, C.A., Hybbinette, S., and Grafstrom, R.C. (1990). Cysteinyl leukotrienes enhance growth of human airway epithelial cells. The American journal of physiology 259, L255-261. 174 Lemaitre, R.N., Rice, K., Marciante, K., Bis, J.C., Lumley, T.S., Wiggins, K.L., Smith, N.L., Heckbert, S.R., and Psaty, B.M. (2009). Variation in eicosanoid genes, non-fatal myocardial infarction and ischemic stroke. Atherosclerosis 204, e58-63. Leppert, D., Hauser, S.L., Kishiyama, J.L., An, S., Zeng, L., and Goetzl, E.J. (1995). Stimulation of matrix metalloproteinase-dependent migration of T cells by eicosanoids. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 9, 14731481. Lewin, A.H., Szewczyk, J., Wilson, J.W., and Carroll, F.I. (2005). Galanthamine analogs: 6Hbenzofuro[3a,3,2,-e,f][1]benzazepine and 6H-benzofuro[3a,3,2-e,f][3]benzazepine. Tetrahedron 61, 7144-7152. Lewis, R.A., Drazen, J.M., Austen, K.F., Clark, D.A., and Corey, E.J. (1980). Identification of the C(6)-S-conjugate of leukotriene A with cysteine as a naturally occurring slow reacting substance of anaphylaxis (SRS-A). Importance of the 11-cis-geometry for biological activity. Biochemical and biophysical research communications 96, 271-277. Ley, K. (1994). Histamine can induce leukocyte rolling in rat mesenteric venules. The American journal of physiology 267, H1017-1023. Li, W., Nava, R.G., Bribriesco, A.C., Zinselmeyer, B.H., Spahn, J.H., Gelman, A.E., Krupnick, A.S., Miller, M.J., and Kreisel, D. (2012). Intravital 2-photon imaging of leukocyte trafficking in beating heart. The Journal of clinical investigation 122, 2499-2508. Linsel-Nitschke, P., Gotz, A., Medack, A., Konig, I.R., Bruse, P., Lieb, W., Mayer, B., Stark, K., Hengstenberg, C., Fischer, M., et al. (2008). Genetic variation in the arachidonate 5lipoxygenase-activating protein (ALOX5AP) is associated with myocardial infarction in the German population. Clinical science 115, 309-315. Liu, M.C., Dube, L.M., and Lancaster, J. (1996). Acute and chronic effects of a 5-lipoxygenase inhibitor in asthma: a 6-month randomized multicenter trial. Zileuton Study Group. The Journal of allergy and clinical immunology 98, 859-871. Lorico, A., Rappa, G., Finch, R.A., Yang, D., Flavell, R.A., and Sartorelli, A.C. (1997). Disruption of the murine MRP (multidrug resistance protein) gene leads to increased sensitivity to etoposide (VP-16) and increased levels of glutathione. Cancer research 57, 5238-5242. Lotzer, K., Jahn, S., Kramer, C., Hildner, M., Nusing, R., Funk, C.D., and Habenicht, A.J. (2007). 5-Lipoxygenase/cyclooxygenase-2 cross-talk through cysteinyl leukotriene receptor 2 in endothelial cells. Prostaglandins & other lipid mediators 84, 108-115. Lotzer, K., Spanbroek, R., Hildner, M., Urbach, A., Heller, R., Bretschneider, E., Galczenski, H., Evans, J.F., and Habenicht, A.J. (2003). Differential leukotriene receptor expression and calcium responses in endothelial cells and macrophages indicate 5-lipoxygenase-dependent circuits of inflammation and atherogenesis. Arterioscler Thromb Vasc Biol 23, e32-36. 175 Luo, M., Jones, S.M., Flamand, N., Aronoff, D.M., Peters-Golden, M., and Brock, T.G. (2005). Phosphorylation by protein kinase a inhibits nuclear import of 5-lipoxygenase. The Journal of biological chemistry 280, 40609-40616. Luo, M., Jones, S.M., Peters-Golden, M., and Brock, T.G. (2003). Nuclear localization of 5lipoxygenase as a determinant of leukotriene B4 synthetic capacity. Proceedings of the National Academy of Sciences of the United States of America 100, 12165-12170. Luo, M., Jones, S.M., Phare, S.M., Coffey, M.J., Peters-Golden, M., and Brock, T.G. (2004). Protein kinase A inhibits leukotriene synthesis by phosphorylation of 5-lipoxygenase on serine 523. The Journal of biological chemistry 279, 41512-41520. Lynch, K.R., O'Neill, G.P., Liu, Q., Im, D.S., Sawyer, N., Metters, K.M., Coulombe, N., Abramovitz, M., Figueroa, D.J., Zeng, Z., et al. (1999). Characterization of the human cysteinyl leukotriene CysLT1 receptor. Nature 399, 789-793. Maclouf, J.A., and Murphy, R.C. (1988). Transcellular metabolism of neutrophil-derived leukotriene A4 by human platelets. A potential cellular source of leukotriene C4. The Journal of biological chemistry 263, 174-181. Maekawa, A., Balestrieri, B., Austen, K.F., and Kanaoka, Y. (2009). GPR17 is a negative regulator of the cysteinyl leukotriene 1 receptor response to leukotriene D4. Proceedings of the National Academy of Sciences of the United States of America 106, 11685-11690. Maekawa, A., Kanaoka, Y., Lam, B.K., and Austen, K.F. (2001). Identification in mice of two isoforms of the cysteinyl leukotriene 1 receptor that result from alternative splicing. Proceedings of the National Academy of Sciences of the United States of America 98, 2256-2261. Maekawa, A., Kanaoka, Y., Xing, W., and Austen, K.F. (2008). Functional recognition of a distinct receptor preferential for leukotriene E4 in mice lacking the cysteinyl leukotriene 1 and 2 receptors. Proceedings of the National Academy of Sciences of the United States of America 105, 16695-16700. Maekawa, A., Xing, W., Austen, K.F., and Kanaoka, Y. (2010). GPR17 regulates immune pulmonary inflammation induced by house dust mites. Journal of immunology 185, 1846-1854. Magnusson, C., Ehrnstrom, R., Olsen, J., and Sjolander, A. (2007). An increased expression of cysteinyl leukotriene 2 receptor in colorectal adenocarcinomas correlates with high differentiation. Cancer research 67, 9190-9198. Magnusson, C., Liu, J., Ehrnstrom, R., Manjer, J., Jirstrom, K., Andersson, T., and Sjolander, A. (2011). Cysteinyl leukotriene receptor expression pattern affects migration of breast cancer cells and survival of breast cancer patients. International journal of cancer Journal international du cancer 129, 9-22. Malmsten, C.L., Palmblad, J., Uden, A.M., Radmark, O., Engstedt, L., and Samuelsson, B. (1980). Leukotriene B4: a highly potent and stereospecific factor stimulating migration of polymorphonuclear leukocytes. Acta physiologica Scandinavica 110, 449-451. 176 Mancuso, P., and Peters-Golden, M. (2000). Modulation of alveolar macrophage phagocytosis by leukotrienes is Fc receptor-mediated and protein kinase C-dependent. American journal of respiratory cell and molecular biology 23, 727-733. Marchant, D.J., Boyd, J.H., Lin, D.C., Granville, D.J., Garmaroudi, F.S., and McManus, B.M. (2012). Inflammation in myocardial diseases. Circulation research 110, 126-144. Martin, V., Sawyer, N., Stocco, R., Unett, D., Lerner, M.R., Abramovitz, M., and Funk, C.D. (2001). Molecular cloning and functional characterization of murine cysteinyl-leukotriene 1 (CysLT(1)) receptors. Biochemical pharmacology 62, 1193-1200. Massoumi, R., and Sjolander, A. (2001). Leukotriene D(4) affects localisation of vinculin in intestinal epithelial cells via distinct tyrosine kinase and protein kinase C controlled events. Journal of cell science 114, 1925-1934. Matsuyama, M., Hayama, T., Funao, K., Kawahito, Y., Sano, H., Takemoto, Y., Nakatani, T., and Yoshimura, R. (2007). Overexpression of cysteinyl LT1 receptor in prostate cancer and CysLT1R antagonist inhibits prostate cancer cell growth through apoptosis. Oncology reports 18, 99-104. Maznyczka, A., Braund, P., Mangino, M., and Samani, N.J. (2008). Arachidonate 5lipoxygenase (5-LO) promoter genotype and risk of myocardial infarction: a case-control study. Atherosclerosis 199, 328-332. McCafferty, D.M., Craig, A.W., Senis, Y.A., and Greer, P.A. (2002). Absence of Fer proteintyrosine kinase exacerbates leukocyte recruitment in response to endotoxin. Journal of immunology 168, 4930-4935. McGee, J.E., and Fitzpatrick, F.A. (1986). Erythrocyte-neutrophil interactions: formation of leukotriene B4 by transcellular biosynthesis. Proceedings of the National Academy of Sciences of the United States of America 83, 1349-1353. McIntyre, T.M., Zimmerman, G.A., and Prescott, S.M. (1986). Leukotrienes C4 and D4 stimulate human endothelial cells to synthesize platelet-activating factor and bind neutrophils. Proceedings of the National Academy of Sciences of the United States of America 83, 22042208. McMahon, B., Stenson, C., McPhillips, F., Fanning, A., Brady, H.R., and Godson, C. (2000). Lipoxin A4 antagonizes the mitogenic effects of leukotriene D4 in human renal mesangial cells. Differential activation of MAP kinases through distinct receptors. The Journal of biological chemistry 275, 27566-27575. McMullen, J.R., and Jennings, G.L. (2007). Differences between pathological and physiological cardiac hypertrophy: novel therapeutic strategies to treat heart failure. Clinical and experimental pharmacology & physiology 34, 255-262. Mellor, E.A., Frank, N., Soler, D., Hodge, M.R., Lora, J.M., Austen, K.F., and Boyce, J.A. (2003). Expression of the type 2 receptor for cysteinyl leukotrienes (CysLT2R) by human mast 177 cells: Functional distinction from CysLT1R. Proceedings of the National Academy of Sciences of the United States of America 100, 11589-11593. Mellor, E.A., Maekawa, A., Austen, K.F., and Boyce, J.A. (2001). Cysteinyl leukotriene receptor 1 is also a pyrimidinergic receptor and is expressed by human mast cells. Proceedings of the National Academy of Sciences of the United States of America 98, 7964-7969. Michelassi, F., Landa, L., Hill, R.D., Lowenstein, E., Watkins, W.D., Petkau, A.J., and Zapol, W.M. (1982). Leukotriene D4: a potent coronary artery vasoconstrictor associated with impaired ventricular contraction. Science 217, 841-843. Mita, H., Hasegawa, M., Saito, H., and Akiyama, K. (2001). Levels of cysteinyl leukotriene receptor mRNA in human peripheral leucocytes: significantly higher expression of cysteinyl leukotriene receptor 2 mRNA in eosinophils. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 31, 1714-1723. Miyahara, N., Takeda, K., Miyahara, S., Matsubara, S., Koya, T., Joetham, A., Krishnan, E., Dakhama, A., Haribabu, B., and Gelfand, E.W. (2005). Requirement for leukotriene B4 receptor 1 in allergen-induced airway hyperresponsiveness. American journal of respiratory and critical care medicine 172, 161-167. Modat, G., Muller, A., Mary, A., Gregoire, C., and Bonne, C. (1987). Differential effects of leukotrienes B4 and C4 on bovine aortic endothelial cell proliferation in vitro. Prostaglandins 33, 531-538. Moos, M.P., and Funk, C.D. (2008). Endothelial cysteinyl leukotriene 2 receptor expression and myocardial ischemia/reperfusion injury. Trends in cardiovascular medicine 18, 268-273. Moos, M.P., Mewburn, J.D., Kan, F.W., Ishii, S., Abe, M., Sakimura, K., Noguchi, K., Shimizu, T., and Funk, C.D. (2008). Cysteinyl leukotriene 2 receptor-mediated vascular permeability via transendothelial vesicle transport. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 22, 4352-4362. Moraes, J., Assreuy, J., Canetti, C., and Barja-Fidalgo, C. (2010). Leukotriene B4 mediates vascular smooth muscle cell migration through alphavbeta3 integrin transactivation. Atherosclerosis 212, 406-413. Moskowitz, M.A., Kiwak, K.J., Hekimian, K., and Levine, L. (1984). Synthesis of compounds with properties of leukotrienes C4 and D4 in gerbil brains after ischemia and reperfusion. Science 224, 886-889. Mosser, D.M., and Edwards, J.P. (2008). Exploring the full spectrum of macrophage activation. Nature reviews Immunology 8, 958-969. Murphy, R.C., and Gijon, M.A. (2007). Biosynthesis and metabolism of leukotrienes. The Biochemical journal 405, 379-395. 178 Murphy, R.C., Hammarstrom, S., and Samuelsson, B. (1979). Leukotriene C: a slow-reacting substance from murine mastocytoma cells. Proceedings of the National Academy of Sciences of the United States of America 76, 4275-4279. Murray, J., Ward, C., O'Flaherty, J.T., Dransfield, I., Haslett, C., Chilvers, E.R., and Rossi, A.G. (2003). Role of leukotrienes in the regulation of human granulocyte behaviour: dissociation between agonist-induced activation and retardation of apoptosis. British journal of pharmacology 139, 388-398. Nabel, E.G., and Braunwald, E. (2012). A tale of coronary artery disease and myocardial infarction. The New England journal of medicine 366, 54-63. Nagy, E., Andersson, D.C., Caidahl, K., Eriksson, M.J., Eriksson, P., Franco-Cereceda, A., Hansson, G.K., and Back, M. (2011). Upregulation of the 5-lipoxygenase pathway in human aortic valves correlates with severity of stenosis and leads to leukotriene-induced effects on valvular myofibroblasts. Circulation 123, 1316-1325. Nahrendorf, M., Pittet, M.J., and Swirski, F.K. (2010). Monocytes: protagonists of infarct inflammation and repair after myocardial infarction. Circulation 121, 2437-2445. Nahrendorf, M., Swirski, F.K., Aikawa, E., Stangenberg, L., Wurdinger, T., Figueiredo, J.L., Libby, P., Weissleder, R., and Pittet, M.J. (2007). The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. The Journal of experimental medicine 204, 3037-3047. National Asthma, E., and Prevention, P. (2007). Expert Panel Report 3 (EPR-3): Guidelines for the Diagnosis and Management of Asthma-Summary Report 2007. The Journal of allergy and clinical immunology 120, S94-138. Ng, C.F., Sun, F.F., Taylor, B.M., Wolin, M.S., and Wong, P.Y. (1991). Functional properties of guinea pig eosinophil leukotriene B4 receptor. Journal of immunology 147, 3096-3103. Ni, N.C., Yan, D., Ballantyne, L.L., Barajas-Espinosa, A., St Amand, T., Pratt, D.A., and Funk, C.D. (2011). A selective cysteinyl leukotriene receptor 2 antagonist blocks myocardial ischemia/reperfusion injury and vascular permeability in mice. The Journal of pharmacology and experimental therapeutics 339, 768-778. Nieves, D., and Moreno, J.J. (2006). Role of 5-lipoxygenase pathway in the regulation of RAW 264.7 macrophage proliferation. Biochemical pharmacology 72, 1022-1030. Nishio, N., Okawa, Y., Sakurai, H., and Isobe, K. (2008). Neutrophil depletion delays wound repair in aged mice. Age 30, 11-19. Nonaka, Y., Hiramoto, T., and Fujita, N. (2005). Identification of endogenous surrogate ligands for human P2Y12 receptors by in silico and in vitro methods. Biochemical and biophysical research communications 337, 281-288. 179 Nothacker, H.P., Wang, Z., Zhu, Y., Reinscheid, R.K., Lin, S.H., and Civelli, O. (2000). Molecular cloning and characterization of a second human cysteinyl leukotriene receptor: discovery of a subtype selective agonist. Molecular pharmacology 58, 1601-1608. O'Hickey, S.P., Arm, J.P., Rees, P.J., Spur, B.W., and Lee, T.H. (1988). The relative responsiveness to inhaled leukotriene E4, methacholine and histamine in normal and asthmatic subjects. The European respiratory journal 1, 913-917. Obata, T., Kobayashi, T., Okada, Y., Nakagawa, N., Terawaki, T., and Aishita, H. (1992). Effect of a peptide leukotriene antagonist, ONO-1078 on antigen-induced airway microvascular leakage in actively sensitized guinea pigs. Life sciences 51, 1577-1583. Ock, S., Ahn, J., Lee, S.H., Park, H., Son, J.W., Oh, J.G., Yang, D.K., Lee, W.S., Kim, H.S., Rho, J., et al. (2012). Receptor activator of nuclear factor-kappaB ligand is a novel inducer of myocardial inflammation. Cardiovascular research 94, 105-114. Ohd, J.F., Nielsen, C.K., Campbell, J., Landberg, G., Lofberg, H., and Sjolander, A. (2003). Expression of the leukotriene D4 receptor CysLT1, COX-2, and other cell survival factors in colorectal adenocarcinomas. Gastroenterology 124, 57-70. Ohd, J.F., Wikstrom, K., and Sjolander, A. (2000). Leukotrienes induce cell-survival signaling in intestinal epithelial cells. Gastroenterology 119, 1007-1018. Okunishi, K., and Peters-Golden, M. (2011). Leukotrienes and airway inflammation. Biochimica et biophysica acta 1810, 1096-1102. Orning, L., Hammarstrom, S., and Samuelsson, B. (1980). Leukotriene D: a slow reacting substance from rat basophilic leukemia cells. Proceedings of the National Academy of Sciences of the United States of America 77, 2014-2017. Palmblad, J., Malmsten, C.L., Uden, A.M., Radmark, O., Engstedt, L., and Samuelsson, B. (1981). Leukotriene B4 is a potent and stereospecific stimulator of neutrophil chemotaxis and adherence. Blood 58, 658-661. Panettieri, R.A., Tan, E.M., Ciocca, V., Luttmann, M.A., Leonard, T.B., and Hay, D.W. (1998). Effects of LTD4 on human airway smooth muscle cell proliferation, matrix expression, and contraction In vitro: differential sensitivity to cysteinyl leukotriene receptor antagonists. American journal of respiratory cell and molecular biology 19, 453-461. Parameswaran, K., Liang, H., Fanat, A., Watson, R., Snider, D.P., and O'Byrne, P.M. (2004). Role for cysteinyl leukotrienes in allergen-induced change in circulating dendritic cell number in asthma. The Journal of allergy and clinical immunology 114, 73-79. Parravicini, C., Abbracchio, M.P., Fantucci, P., and Ranghino, G. (2010). Forced unbinding of GPR17 ligands from wild type and R255I mutant receptor models through a computational approach. BMC structural biology 10, 8. 180 Parravicini, C., Ranghino, G., Abbracchio, M.P., and Fantucci, P. (2008). GPR17: molecular modeling and dynamics studies of the 3-D structure and purinergic ligand binding features in comparison with P2Y receptors. BMC bioinformatics 9, 263. Paruchuri, S., Hallberg, B., Juhas, M., Larsson, C., and Sjolander, A. (2002). Leukotriene D(4) activates MAPK through a Ras-independent but PKCepsilon-dependent pathway in intestinal epithelial cells. Journal of cell science 115, 1883-1893. Paruchuri, S., Jiang, Y., Feng, C., Francis, S.A., Plutzky, J., and Boyce, J.A. (2008). Leukotriene E4 activates peroxisome proliferator-activated receptor gamma and induces prostaglandin D2 generation by human mast cells. The Journal of biological chemistry 283, 16477-16487. Paruchuri, S., and Sjolander, A. (2003). Leukotriene D4 mediates survival and proliferation via separate but parallel pathways in the human intestinal epithelial cell line Int 407. The Journal of biological chemistry 278, 45577-45585. Paruchuri, S., Tashimo, H., Feng, C., Maekawa, A., Xing, W., Jiang, Y., Kanaoka, Y., Conley, P., and Boyce, J.A. (2009). Leukotriene E4-induced pulmonary inflammation is mediated by the P2Y12 receptor. The Journal of experimental medicine 206, 2543-2555. Patel, A.S., Smith, A., Nucera, S., Biziato, D., Saha, P., Attia, R.Q., Humphries, J., Mattock, K., Grover, S.P., Lyons, O.T., et al. (2013). TIE2-expressing monocytes/macrophages regulate revascularization of the ischemic limb. EMBO molecular medicine. Patten, R.D., Aronovitz, M.J., Deras-Mejia, L., Pandian, N.G., Hanak, G.G., Smith, J.J., Mendelsohn, M.E., and Konstam, M.A. (1998). Ventricular remodeling in a mouse model of myocardial infarction. The American journal of physiology 274, H1812-1820. Pedersen, K.E., Bochner, B.S., and Undem, B.J. (1997). Cysteinyl leukotrienes induce P-selectin expression in human endothelial cells via a non-CysLT1 receptor-mediated mechanism. The Journal of pharmacology and experimental therapeutics 281, 655-662. Penrose, J.F. (1999). LTC4 synthase. Enzymology, biochemistry, and molecular characterization. Clinical reviews in allergy & immunology 17, 133-152. Poeckel, D., and Funk, C.D. (2010). The 5-lipoxygenase/leukotriene pathway in preclinical models of cardiovascular disease. Cardiovascular research 86, 243-253. Pouliot, M., McDonald, P.P., Borgeat, P., and McColl, S.R. (1994). Granulocyte/macrophage colony-stimulating factor stimulates the expression of the 5-lipoxygenase-activating protein (FLAP) in human neutrophils. The Journal of experimental medicine 179, 1225-1232. Pueringer, R.J., Bahns, C.C., and Hunninghake, G.W. (1992). Alveolar macrophages have greater amounts of the enzyme 5-lipoxygenase than do monocytes. Journal of applied physiology 73, 781-786. Qi, A.D., Harden, T.K., and Nicholas, R.A. (2004). GPR80/99, proposed to be the P2Y(15) receptor activated by adenosine and AMP, is not a P2Y receptor. Purinergic signalling 1, 67-74. 181 Qi, L.L., Fang, S.H., Shi, W.Z., Huang, X.Q., Zhang, X.Y., Lu, Y.B., Zhang, W.P., and Wei, E.Q. (2011). CysLT2 receptor-mediated AQP4 up-regulation is involved in ischemic-like injury through activation of ERK and p38 MAPK in rat astrocytes. Life sciences 88, 50-56. Qiu, H., Gabrielsen, A., Agardh, H.E., Wan, M., Wetterholm, A., Wong, C.H., Hedin, U., Swedenborg, J., Hansson, G.K., Samuelsson, B., et al. (2006a). Expression of 5-lipoxygenase and leukotriene A4 hydrolase in human atherosclerotic lesions correlates with symptoms of plaque instability. Proceedings of the National Academy of Sciences of the United States of America 103, 8161-8166. Qiu, H., Johansson, A.S., Sjostrom, M., Wan, M., Schroder, O., Palmblad, J., and Haeggstrom, J.Z. (2006b). Differential induction of BLT receptor expression on human endothelial cells by lipopolysaccharide, cytokines, and leukotriene B4. Proceedings of the National Academy of Sciences of the United States of America 103, 6913-6918. Radmark, O., and Samuelsson, B. (2007). 5-lipoxygenase: regulation and possible involvement in atherosclerosis. Prostaglandins & other lipid mediators 83, 162-174. Radmark, O., Werz, O., Steinhilber, D., and Samuelsson, B. (2007). 5-Lipoxygenase: regulation of expression and enzyme activity. Trends in biochemical sciences 32, 332-341. Raedschelders, K., Ansley, D.M., and Chen, D.D. (2012). The cellular and molecular origin of reactive oxygen species generation during myocardial ischemia and reperfusion. Pharmacology & therapeutics 133, 230-255. Rao, A.M., Hatcher, J.F., Kindy, M.S., and Dempsey, R.J. (1999). Arachidonic acid and leukotriene C4: role in transient cerebral ischemia of gerbils. Neurochemical research 24, 12251232. Rao, R.M., Yang, L., Garcia-Cardena, G., and Luscinskas, F.W. (2007). Endothelial-dependent mechanisms of leukocyte recruitment to the vascular wall. Circulation research 101, 234-247. Reddy, K.V., Serio, K.J., Hodulik, C.R., and Bigby, T.D. (2003). 5-lipoxygenase-activating protein gene expression. Key role of CCAAT/enhancer-binding proteins (C/EBP) in constitutive and tumor necrosis factor (TNF) alpha-induced expression in THP-1 cells. The Journal of biological chemistry 278, 13810-13818. Reiss, T.F., Chervinsky, P., Dockhorn, R.J., Shingo, S., Seidenberg, B., and Edwards, T.B. (1998). Montelukast, a once-daily leukotriene receptor antagonist, in the treatment of chronic asthma: a multicenter, randomized, double-blind trial. Montelukast Clinical Research Study Group. Archives of internal medicine 158, 1213-1220. Rioux, N., and Castonguay, A. (1998). Inhibitors of lipoxygenase: a new class of cancer chemopreventive agents. Carcinogenesis 19, 1393-1400. Rius, M., Hummel-Eisenbeiss, J., and Keppler, D. (2008). ATP-dependent transport of leukotrienes B4 and C4 by the multidrug resistance protein ABCC4 (MRP4). The Journal of pharmacology and experimental therapeutics 324, 86-94. 182 Roger, V.L., Go, A.S., Lloyd-Jones, D.M., Benjamin, E.J., Berry, J.D., Borden, W.B., Bravata, D.M., Dai, S., Ford, E.S., Fox, C.S., et al. (2012). Heart disease and stroke statistics--2012 update: a report from the American Heart Association. Circulation 125, e2-e220. Rola-Pleszczynski, M., Chavaillaz, P.A., and Lemaire, I. (1986). Stimulation of interleukin 2 and interferon gamma production by leukotriene B4 in human lymphocyte cultures. Prostaglandins, leukotrienes, and medicine 23, 207-210. Rola-Pleszczynski, M., and Lemaire, I. (1985). Leukotrienes augment interleukin 1 production by human monocytes. Journal of immunology 135, 3958-3961. Rola-Pleszczynski, M., and Stankova, J. (1992). Leukotriene B4 enhances interleukin-6 (IL-6) production and IL-6 messenger RNA accumulation in human monocytes in vitro: transcriptional and posttranscriptional mechanisms. Blood 80, 1004-1011. Romson, J.L., Hook, B.G., Kunkel, S.L., Abrams, G.D., Schork, M.A., and Lucchesi, B.R. (1983). Reduction of the extent of ischemic myocardial injury by neutrophil depletion in the dog. Circulation 67, 1016-1023. Rouzer, C.A., and Samuelsson, B. (1985). On the nature of the 5-lipoxygenase reaction in human leukocytes: enzyme purification and requirement for multiple stimulatory factors. Proceedings of the National Academy of Sciences of the United States of America 82, 6040-6044. Rouzer, C.A., Shimizu, T., and Samuelsson, B. (1985). On the nature of the 5-lipoxygenase reaction in human leukocytes: characterization of a membrane-associated stimulatory factor. Proceedings of the National Academy of Sciences of the United States of America 82, 75057509. Rovetto, M.J., Lamberton, W.F., and Neely, J.R. (1975). Mechanisms of glycolytic inhibition in ischemic rat hearts. Circulation research 37, 742-751. Sakata, K., Dahlen, S.E., and Back, M. (2004). The contractile action of leukotriene B4 in the guinea-pig lung involves a vascular component. British journal of pharmacology 141, 449-456. Sala, A., Bolla, M., Zarini, S., Muller-Peddinghaus, R., and Folco, G. (1996). Release of leukotriene A4 versus leukotriene B4 from human polymorphonuclear leukocytes. The Journal of biological chemistry 271, 17944-17948. Sala, A., Rossoni, G., Berti, F., Buccellati, C., Bonazzi, A., Maclouf, J., and Folco, G. (2000). Monoclonal anti-CD18 antibody prevents transcellular biosynthesis of cysteinyl leukotrienes in vitro and in vivo and protects against leukotriene-dependent increase in coronary vascular resistance and myocardial stiffness. Circulation 101, 1436-1440. Sala, A., Rossoni, G., Buccellati, C., Berti, F., Folco, G., and Maclouf, J. (1993). Formation of sulphidopeptide-leukotrienes by cell-cell interaction causes coronary vasoconstriction in isolated, cell-perfused heart of rabbit. British journal of pharmacology 110, 1206-1212. 183 Samuelsson, B. (1983). Leukotrienes: mediators of immediate hypersensitivity reactions and inflammation. Science 220, 568-575. Sarau, H.M., Ames, R.S., Chambers, J., Ellis, C., Elshourbagy, N., Foley, J.J., Schmidt, D.B., Muccitelli, R.M., Jenkins, O., Murdock, P.R., et al. (1999). Identification, molecular cloning, expression, and characterization of a cysteinyl leukotriene receptor. Molecular pharmacology 56, 657-663. Schafer, C., Ladilov, Y., Inserte, J., Schafer, M., Haffner, S., Garcia-Dorado, D., and Piper, H.M. (2001). Role of the reverse mode of the Na+/Ca2+ exchanger in reoxygenation-induced cardiomyocyte injury. Cardiovascular research 51, 241-250. Schellenberg, R.R., and Foster, A. (1984). Differential activity of leukotrienes upon human pulmonary vein and artery. Prostaglandins 27, 475-482. Schievella, A.R., Regier, M.K., Smith, W.L., and Lin, L.L. (1995). Calcium-mediated translocation of cytosolic phospholipase A2 to the nuclear envelope and endoplasmic reticulum. The Journal of biological chemistry 270, 30749-30754. Schroder, O., Sjostrom, M., Qiu, H., Jakobsson, P.J., and Haeggstrom, J.Z. (2005). Microsomal glutathione S-transferases: selective up-regulation of leukotriene C4 synthase during lipopolysaccharide-induced pyresis. Cellular and molecular life sciences : CMLS 62, 87-94. Scoggan, K.A., Jakobsson, P.J., and Ford-Hutchinson, A.W. (1997). Production of leukotriene C4 in different human tissues is attributable to distinct membrane bound biosynthetic enzymes. The Journal of biological chemistry 272, 10182-10187. Serhan, C.N., Radin, A., Smolen, J.E., Korchak, H., Samuelsson, B., and Weissmann, G. (1982). Leukotriene B4 is a complete secretagogue in human neutrophils: a kinetic analysis. Biochemical and biophysical research communications 107, 1006-1012. Sharma, V., Dadheech, S., Kaul, S., Jyothy, A., and Munshi, A. (2013). Association of ALOX5AP1 SG13S114T/A variant with ischemic stroke, stroke subtypes and aspirin resistance. Journal of the neurological sciences 331, 108-113. Sheng, W.W., Li, C.T., Zhang, W.P., Yuan, Y.M., Hu, H., Fang, S.H., Zhang, L., and Wei, E.Q. (2006). Distinct roles of CysLT1 and CysLT2 receptors in oxygen glucose deprivation-induced PC12 cell death. Biochemical and biophysical research communications 346, 19-25. Shi, Q.J., Xiao, L., Zhao, B., Zhang, X.Y., Wang, X.R., Xu, D.M., Yu, S.Y., Fang, S.H., Lu, Y.B., Zhang, W.P., et al. (2012a). Intracerebroventricular injection of HAMI 3379, a selective cysteinyl leukotriene receptor 2 antagonist, protects against acute brain injury after focal cerebral ischemia in rats. Brain research 1484, 57-67. Shi, W.Z., Zhao, C.Z., Zhao, B., Shi, Q.J., Zhang, L.H., Wang, Y.F., Fang, S.H., Lu, Y.B., Zhang, W.P., and Wei, E.Q. (2012b). Aggravated inflammation and increased expression of cysteinyl leukotriene receptors in the brain after focal cerebral ischemia in AQP4-deficient mice. Neuroscience bulletin 28, 680-692. 184 Shin, E.H., Lee, H.Y., and Bae, Y.S. (2006). Leukotriene B4 stimulates human monocytederived dendritic cell chemotaxis. Biochemical and biophysical research communications 348, 606-611. Shirasaki, H., Seki, N., Fujita, M., Kikuchi, M., Kanaizumi, E., Watanabe, K., and Himi, T. (2007). Agonist- and T(H)2 cytokine-induced up-regulation of cysteinyl leukotriene receptor messenger RNA in human monocytes. Annals of allergy, asthma & immunology : official publication of the American College of Allergy, Asthma, & Immunology 99, 340-347. Shishido, T., Nozaki, N., Yamaguchi, S., Shibata, Y., Nitobe, J., Miyamoto, T., Takahashi, H., Arimoto, T., Maeda, K., Yamakawa, M., et al. (2003). Toll-like receptor-2 modulates ventricular remodeling after myocardial infarction. Circulation 108, 2905-2910. Sjostrom, M., Jakobsson, P.J., Heimburger, M., Palmblad, J., and Haeggstrom, J.Z. (2001). Human umbilical vein endothelial cells generate leukotriene C4 via microsomal glutathione Stransferase type 2 and express the CysLT(1) receptor. European journal of biochemistry / FEBS 268, 2578-2586. Sjostrom, M., Johansson, A.S., Schroder, O., Qiu, H., Palmblad, J., and Haeggstrom, J.Z. (2003). Dominant expression of the CysLT2 receptor accounts for calcium signaling by cysteinyl leukotrienes in human umbilical vein endothelial cells. Arteriosclerosis, thrombosis, and vascular biology 23, e37-41. Smith, S.C., Jr., Allen, J., Blair, S.N., Bonow, R.O., Brass, L.M., Fonarow, G.C., Grundy, S.M., Hiratzka, L., Jones, D., Krumholz, H.M., et al. (2006). AHA/ACC guidelines for secondary prevention for patients with coronary and other atherosclerotic vascular disease: 2006 update: endorsed by the National Heart, Lung, and Blood Institute. Circulation 113, 2363-2372. Smyth, E.M., Grosser, T., Wang, M., Yu, Y., and FitzGerald, G.A. (2009). Prostanoids in health and disease. Journal of lipid research 50 Suppl, S423-428. Song, K., Nam, Y.J., Luo, X., Qi, X., Tan, W., Huang, G.N., Acharya, A., Smith, C.L., Tallquist, M.D., Neilson, E.G., et al. (2012). Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 485, 599-604. Soter, N.A., Lewis, R.A., Corey, E.J., and Austen, K.F. (1983). Local effects of synthetic leukotrienes (LTC4, LTD4, LTE4, and LTB4) in human skin. The Journal of investigative dermatology 80, 115-119. Spanbroek, R., Grabner, R., Lotzer, K., Hildner, M., Urbach, A., Ruhling, K., Moos, M.P., Kaiser, B., Cohnert, T.U., Wahlers, T., et al. (2003). Expanding expression of the 5-lipoxygenase pathway within the arterial wall during human atherogenesis. Proceedings of the National Academy of Sciences of the United States of America 100, 1238-1243. Steffens, S., Montecucco, F., and Mach, F. (2009). The inflammatory response as a target to reduce myocardial ischaemia and reperfusion injury. Thrombosis and haemostasis 102, 240-247. 185 Suissa, S., Dennis, R., Ernst, P., Sheehy, O., and Wood-Dauphinee, S. (1997). Effectiveness of the leukotriene receptor antagonist zafirlukast for mild-to-moderate asthma. A randomized, double-blind, placebo-controlled trial. Annals of internal medicine 126, 177-183. Tada, M., Kuzuya, T., Hoshida, S., and Nishida, M. (1988). Arachidonate metabolism in myocardial ischemia and reperfusion. Journal of molecular and cellular cardiology 20 Suppl 2, 135-143. Takasaki, J., Kamohara, M., Matsumoto, M., Saito, T., Sugimoto, T., Ohishi, T., Ishii, H., Ota, T., Nishikawa, T., Kawai, Y., et al. (2000). The molecular characterization and tissue distribution of the human cysteinyl leukotriene CysLT(2) receptor. Biochemical and biophysical research communications 274, 316-322. Takase, B., Kurita, A., Maruyama, T., Uehata, A., Nishioka, T., Mizuno, K., Nakamura, H., Katsura, K., and Kanda, Y. (1996). Change of plasma leukotriene C4 during myocardial ischemia in humans. Clinical cardiology 19, 198-204. Takemura, G., Nakagawa, M., Kanamori, H., Minatoguchi, S., and Fujiwara, H. (2009). Benefits of reperfusion beyond infarct size limitation. Cardiovascular research 83, 269-276. Tanimoto, Y., Takahashi, K., and Kimura, I. (1992). Effects of cytokines on human basophil chemotaxis. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 22, 1020-1025. Taylor, I.K., O'Shaughnessy, K.M., Fuller, R.W., and Dollery, C.T. (1991). Effect of cysteinylleukotriene receptor antagonist ICI 204.219 on allergen-induced bronchoconstriction and airway hyperreactivity in atopic subjects. Lancet 337, 690-694. Taylor, I.K., Wellings, R., Taylor, G.W., and Fuller, R.W. (1992). Urinary leukotriene E4 excretion in exercise-induced asthma. Journal of applied physiology 73, 743-748. Terashima, T., Amakawa, K., Matsumaru, A., and Yamaguchi, K. (2002). Correlation between cysteinyl leukotriene release from leukocytes and clinical response to a leukotriene inhibitor. Chest 122, 1566-1570. Terawaki, K., Yokomizo, T., Nagase, T., Toda, A., Taniguchi, M., Hashizume, K., Yagi, T., and Shimizu, T. (2005). Absence of leukotriene B4 receptor 1 confers resistance to airway hyperresponsiveness and Th2-type immune responses. Journal of immunology 175, 4217-4225. Thivierge, M., Doty, M., Johnson, J., Stankova, J., and Rola-Pleszczynski, M. (2000). IL-5 upregulates cysteinyl leukotriene 1 receptor expression in HL-60 cells differentiated into eosinophils. Journal of immunology 165, 5221-5226. Thivierge, M., Stankova, J., and Rola-Pleszczynski, M. (2001). IL-13 and IL-4 up-regulate cysteinyl leukotriene 1 receptor expression in human monocytes and macrophages. Journal of immunology 167, 2855-2860. 186 Thivierge, M., Stankova, J., and Rola-Pleszczynski, M. (2006). Toll-like receptor agonists differentially regulate cysteinyl-leukotriene receptor 1 expression and function in human dendritic cells. The Journal of allergy and clinical immunology 117, 1155-1162. Thivierge, M., Stankova, J., and Rola-Pleszczynski, M. (2009). Cysteinyl-leukotriene receptor type 1 expression and function is down-regulated during monocyte-derived dendritic cell maturation with zymosan: involvement of IL-10 and prostaglandins. Journal of immunology 183, 6778-6787. Thompson, C., Cloutier, A., Bosse, Y., Poisson, C., Larivee, P., McDonald, P.P., Stankova, J., and Rola-Pleszczynski, M. (2008). Signaling by the cysteinyl-leukotriene receptor 2. Involvement in chemokine gene transcription. The Journal of biological chemistry 283, 19741984. Thompson, C., Cloutier, A., Bosse, Y., Thivierge, M., Gouill, C.L., Larivee, P., McDonald, P.P., Stankova, J., and Rola-Pleszczynski, M. (2006). CysLT1 receptor engagement induces activator protein-1- and NF-kappaB-dependent IL-8 expression. American journal of respiratory cell and molecular biology 35, 697-704. Toki, Y., Hieda, N., Torii, T., Hashimoto, H., Ito, T., Ogawa, K., and Satake, T. (1988). The effects of lipoxygenase inhibitor and peptidoleukotriene antagonist on myocardial injury in a canine coronary occlusion-reperfusion model. Prostaglandins 35, 555-571. Toldo, S., Breckenridge, D.G., Mezzaroma, E., Van Tassell, B.W., Shryock, J., Kannan, H., Phan, D., Budas, G., Farkas, D., Lesnefsky, E., et al. (2012). Inhibition of apoptosis signalregulating kinase 1 reduces myocardial ischemia-reperfusion injury in the mouse. Journal of the American Heart Association 1, e002360. Tong, W.G., Ding, X.Z., Talamonti, M.S., Bell, R.H., and Adrian, T.E. (2005). LTB4 stimulates growth of human pancreatic cancer cells via MAPK and PI-3 kinase pathways. Biochemical and biophysical research communications 335, 949-956. Tsopanoglou, N.E., Pipili-Synetos, E., and Maragoudakis, M.E. (1994). Leukotrienes C4 and D4 promote angiogenesis via a receptor-mediated interaction. European journal of pharmacology 258, 151-154. Tudhope, S.R., Cuthbert, N.J., Abram, T.S., Jennings, M.A., Maxey, R.J., Thompson, A.M., Norman, P., and Gardiner, P.J. (1994). BAY u9773, a novel antagonist of cysteinyl-leukotrienes with activity against two receptor subtypes. European journal of pharmacology 264, 317-323. Ueda, T., Takeno, S., Furukido, K., Hirakawa, K., and Yajin, K. (2003). Leukotriene receptor antagonist pranlukast suppresses eosinophil infiltration and cytokine production in human nasal mucosa of perennial allergic rhinitis. The Annals of otology, rhinology, and laryngology 112, 955-961. Uzonyi, B., Lotzer, K., Jahn, S., Kramer, C., Hildner, M., Bretschneider, E., Radke, D., Beer, M., Vollandt, R., Evans, J.F., et al. (2006). Cysteinyl leukotriene 2 receptor and protease-activated 187 receptor 1 activate strongly correlated early genes in human endothelial cells. Proceedings of the National Academy of Sciences of the United States of America 103, 6326-6331. Vaddi, K., and Newton, R.C. (1994). Regulation of monocyte integrin expression by beta-family chemokines. Journal of immunology 153, 4721-4732. Vakeva, A.P., Agah, A., Rollins, S.A., Matis, L.A., Li, L., and Stahl, G.L. (1998). Myocardial infarction and apoptosis after myocardial ischemia and reperfusion: role of the terminal complement components and inhibition by anti-C5 therapy. Circulation 97, 2259-2267. van Amerongen, M.J., Harmsen, M.C., van Rooijen, N., Petersen, A.H., and van Luyn, M.J. (2007). Macrophage depletion impairs wound healing and increases left ventricular remodeling after myocardial injury in mice. The American journal of pathology 170, 818-829. Van de Werf, F., Bax, J., Betriu, A., Blomstrom-Lundqvist, C., Crea, F., Falk, V., Filippatos, G., Fox, K., Huber, K., Kastrati, A., et al. (2008). Management of acute myocardial infarction in patients presenting with persistent ST-segment elevation: the Task Force on the Management of ST-Segment Elevation Acute Myocardial Infarction of the European Society of Cardiology. European heart journal 29, 2909-2945. Vandervelde, S., van Luyn, M.J., Rozenbaum, M.H., Petersen, A.H., Tio, R.A., and Harmsen, M.C. (2007). Stem cell-related cardiac gene expression early after murine myocardial infarction. Cardiovascular research 73, 783-793. Vanhoutte, D., Schellings, M.W., Gotte, M., Swinnen, M., Herias, V., Wild, M.K., Vestweber, D., Chorianopoulos, E., Cortes, V., Rigotti, A., et al. (2007). Increased expression of syndecan-1 protects against cardiac dilatation and dysfunction after myocardial infarction. Circulation 115, 475-482. Vannella, K.M., McMillan, T.R., Charbeneau, R.P., Wilke, C.A., Thomas, P.E., Toews, G.B., Peters-Golden, M., and Moore, B.B. (2007). Cysteinyl leukotrienes are autocrine and paracrine regulators of fibrocyte function. J Immunol 179, 7883-7890. Venneri, M.A., De Palma, M., Ponzoni, M., Pucci, F., Scielzo, C., Zonari, E., Mazzieri, R., Doglioni, C., and Naldini, L. (2007). Identification of proangiogenic TIE2-expressing monocytes (TEMs) in human peripheral blood and cancer. Blood 109, 5276-5285. Vinten-Johansen, J. (2004). Involvement of neutrophils in the pathogenesis of lethal myocardial reperfusion injury. Cardiovascular research 61, 481-497. Voelkel, N.F., Czartolomna, J., Simpson, J., and Murphy, R.C. (1992). FMLP causes eicosanoiddependent vasoconstriction and edema in lungs from endotoxin-primed rats. The American review of respiratory disease 145, 701-711. Wada, K., Arita, M., Nakajima, A., Katayama, K., Kudo, C., Kamisaki, Y., and Serhan, C.N. (2006). Leukotriene B4 and lipoxin A4 are regulatory signals for neural stem cell proliferation and differentiation. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 20, 1785-1792. 188 Wang, G., Zhang, J., Sun, H., Cao, W., Zhang, J., Wang, Y., and Xiao, H. (2012). Genetic variation in members of the leukotrienes biosynthesis pathway confers risk of ischemic stroke in Eastern Han Chinese. Prostaglandins, leukotrienes, and essential fatty acids 87, 169-175. Wang, P., Chen, H., Qin, H., Sankarapandi, S., Becher, M.W., Wong, P.C., and Zweier, J.L. (1998). Overexpression of human copper, zinc-superoxide dismutase (SOD1) prevents postischemic injury. Proceedings of the National Academy of Sciences of the United States of America 95, 4556-4560. Weiss, J.W., Drazen, J.M., Coles, N., McFadden, E.R., Jr., Weller, P.F., Corey, E.J., Lewis, R.A., and Austen, K.F. (1982). Bronchoconstrictor effects of leukotriene C in humans. Science 216, 196-198. Weller, C.L., Collington, S.J., Brown, J.K., Miller, H.R., Al-Kashi, A., Clark, P., Jose, P.J., Hartnell, A., and Williams, T.J. (2005). Leukotriene B4, an activation product of mast cells, is a chemoattractant for their progenitors. The Journal of experimental medicine 201, 1961-1971. Wenzel, S., Busse, W., Calhoun, W., Panettieri, R., Jr., Peters-Golden, M., Dube, L., WaltonBowen, K., Russell, H., and Harris, J. (2007). The safety and efficacy of zileuton controlledrelease tablets as adjunctive therapy to usual care in the treatment of moderate persistent asthma: a 6-month randomized controlled study. The Journal of asthma : official journal of the Association for the Care of Asthma 44, 305-310. Wenzel, S.E. (2003). The role of leukotrienes in asthma. Prostaglandins, leukotrienes, and essential fatty acids 69, 145-155. Wenzel, S.E., Larsen, G.L., Johnston, K., Voelkel, N.F., and Westcott, J.Y. (1990). Elevated levels of leukotriene C4 in bronchoalveolar lavage fluid from atopic asthmatics after endobronchial allergen challenge. The American review of respiratory disease 142, 112-119. Wenzel, S.E., Trudeau, J.B., Kaminsky, D.A., Cohn, J., Martin, R.J., and Westcott, J.Y. (1995). Effect of 5-lipoxygenase inhibition on bronchoconstriction and airway inflammation in nocturnal asthma. American journal of respiratory and critical care medicine 152, 897-905. Werz, O., Burkert, E., Fischer, L., Szellas, D., Dishart, D., Samuelsson, B., Radmark, O., and Steinhilber, D. (2002). Extracellular signal-regulated kinases phosphorylate 5-lipoxygenase and stimulate 5-lipoxygenase product formation in leukocytes. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 16, 1441-1443. Werz, O., Klemm, J., Samuelsson, B., and Radmark, O. (2000). 5-lipoxygenase is phosphorylated by p38 kinase-dependent MAPKAP kinases. Proceedings of the National Academy of Sciences of the United States of America 97, 5261-5266. Westerberg, E., Deshpande, J.K., and Wieloch, T. (1987). Regional differences in arachidonic acid release in rat hippocampal CA1 and CA3 regions during cerebral ischemia. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 7, 189-192. 189 White, H.D., and Chew, D.P. (2008). Acute myocardial infarction. Lancet 372, 570-584. Wikstrom, K., Ohd, J.F., and Sjolander, A. (2003). Regulation of leukotriene-dependent induction of cyclooxygenase-2 and Bcl-2. Biochemical and biophysical research communications 302, 330-335. Woods, J.W., Coffey, M.J., Brock, T.G., Singer, II, and Peters-Golden, M. (1995). 5Lipoxygenase is located in the euchromatin of the nucleus in resting human alveolar macrophages and translocates to the nuclear envelope upon cell activation. The Journal of clinical investigation 95, 2035-2046. Woszczek, G., Chen, L.Y., Alsaaty, S., Nagineni, S., and Shelhamer, J.H. (2010). Concentrationdependent noncysteinyl leukotriene type 1 receptor-mediated inhibitory activity of leukotriene receptor antagonists. Journal of immunology 184, 2219-2225. Woszczek, G., Chen, L.Y., Nagineni, S., Alsaaty, S., Harry, A., Logun, C., Pawliczak, R., and Shelhamer, J.H. (2007). IFN-gamma induces cysteinyl leukotriene receptor 2 expression and enhances the responsiveness of human endothelial cells to cysteinyl leukotrienes. Journal of immunology 178, 5262-5270. Woszczek, G., Chen, L.Y., Nagineni, S., and Shelhamer, J.H. (2008). IL-10 inhibits cysteinyl leukotriene-induced activation of human monocytes and monocyte-derived dendritic cells. Journal of immunology 180, 7597-7603. Woszczek, G., Pawliczak, R., Qi, H.Y., Nagineni, S., Alsaaty, S., Logun, C., and Shelhamer, J.H. (2005). Functional characterization of human cysteinyl leukotriene 1 receptor gene structure. Journal of immunology 175, 5152-5159. Wunder, F., Tinel, H., Kast, R., Geerts, A., Becker, E.M., Kolkhof, P., Hutter, J., Erguden, J., and Harter, M. (2010). Pharmacological characterization of the first potent and selective antagonist at the cysteinyl leukotriene 2 (CysLT(2)) receptor. British journal of pharmacology 160, 399-409. Yamaoka, K.A., and Kolb, J.P. (1993). Leukotriene B4 induces interleukin 5 generation from human T lymphocytes. European journal of immunology 23, 2392-2398. Yan, D., Stocco, R., Sawyer, N., Nesheim, M.E., Abramovitz, M., and Funk, C.D. (2011). Differential signaling of cysteinyl leukotrienes and a novel cysteinyl leukotriene receptor 2 (CysLT(2)) agonist, N-methyl-leukotriene C(4), in calcium reporter and beta arrestin assays. Molecular pharmacology 79, 270-278. Yang, F., Liu, Y.H., Yang, X.P., Xu, J., Kapke, A., and Carretero, O.A. (2002). Myocardial infarction and cardiac remodelling in mice. Exp Physiol 87, 547-555. Yasojima, K., Schwab, C., McGeer, E.G., and McGeer, P.L. (1998). Human heart generates complement proteins that are upregulated and activated after myocardial infarction. Circulation research 83, 860-869. 190 Ye, Y.N., Wu, W.K., Shin, V.Y., Bruce, I.C., Wong, B.C., and Cho, C.H. (2005). Dual inhibition of 5-LOX and COX-2 suppresses colon cancer formation promoted by cigarette smoke. Carcinogenesis 26, 827-834. Yellon, D.M., and Hausenloy, D.J. (2007). Myocardial reperfusion injury. The New England journal of medicine 357, 1121-1135. Zarini, S., Gijon, M.A., Ransome, A.E., Murphy, R.C., and Sala, A. (2009). Transcellular biosynthesis of cysteinyl leukotrienes in vivo during mouse peritoneal inflammation. Proceedings of the National Academy of Sciences of the United States of America 106, 82968301. Zhang, L.Z., and Guo, Z. (2009). Tramadol reduces myocardial infarct size and expression and activation of nuclear factor kappa B in acute myocardial infarction in rats. European journal of anaesthesiology 26, 1048-1055. Zhang, R., Khoo, M.S., Wu, Y., Yang, Y., Grueter, C.E., Ni, G., Price, E.E., Jr., Thiel, W., Guatimosim, S., Song, L.S., et al. (2005). Calmodulin kinase II inhibition protects against structural heart disease. Nature medicine 11, 409-417. Zhang, W.P., Hu, H., Zhang, L., Ding, W., Yao, H.T., Chen, K.D., Sheng, W.W., Chen, Z., and Wei, E.Q. (2004). Expression of cysteinyl leukotriene receptor 1 in human traumatic brain injury and brain tumors. Neuroscience letters 363, 247-251. Zhang, W.P., Wei, E.Q., Mei, R.H., Zhu, C.Y., and Zhao, M.H. (2002). Neuroprotective effect of ONO-1078, a leukotriene receptor antagonist, on focal cerebral ischemia in rats. Acta pharmacologica Sinica 23, 871-877. Zhang, X.Y., Wang, X.R., Xu, D.M., Yu, S.Y., Shi, Q.J., Zhang, L.H., Chen, L., Fang, S.H., Lu, Y.B., Zhang, W.P., et al. (2013). HAMI 3379, a CysLT2 receptor antagonist, attenuates ischemia-like neuronal injury by inhibiting microglial activation. The Journal of pharmacology and experimental therapeutics. Zhao, C.Z., Zhao, B., Zhang, X.Y., Huang, X.Q., Shi, W.Z., Liu, H.L., Fang, S.H., Lu, Y.B., Zhang, W.P., Tang, F.D., et al. (2011). Cysteinyl leukotriene receptor 2 is spatiotemporally involved in neuron injury, astrocytosis and microgliosis after focal cerebral ischemia in rats. Neuroscience 189, 1-11. Zhou, G., Zhou, Z., Ge, S., Liu, D., Zhang, R., Xu, G., Zhu, W., Yin, Q., Chen, A.F., and Liu, X. (2009). IL-18 accelerates the cell apoptosis by up-regulating cysteinyl leukotriene 2 receptor expression in human umbilical vein endothelial cells at the early stage of administration. Vascular pharmacology 50, 171-177. Ziegler-Heitbrock, L. (2007). The CD14+ CD16+ blood monocytes: their role in infection and inflammation. Journal of leukocyte biology 81, 584-592. 191 Zintzaras, E., Rodopoulou, P., and Sakellaridis, N. (2009). Variants of the arachidonate 5lipoxygenase-activating protein (ALOX5AP) gene and risk of stroke: a HuGE gene-disease association review and meta-analysis. American journal of epidemiology 169, 523-532. Zweier, J.L. (1988). Measurement of superoxide-derived free radicals in the reperfused heart. Evidence for a free radical mechanism of reperfusion injury. The Journal of biological chemistry 263, 1353-1357. 192 Appendix Formulae Used to Calculate Cardiac Function from Echocardiography Images Left ventricular volume, diastole (LV Vol;d) = ((7.0 / (2.4 + LVID;d)) * LVID;d3 P Left ventricular volume, systole (LV Vol;s) = ((7.0 / (2.4 + LVID;s)) * LVID;s3 P Ejection fraction = 100 * ((LV Vol;d – LV Vol;s) / LV Vol;d) Fractional shortening = 100 * ((LVID;d – LVID;s) / LVID;d) Stroke volume (SV) = LV Vol;d – LV Vol;s Cardiac output = SV/HR Left ventricular mass = 1.053 * ((LVID;d + LVPW;d + IVS;d)3– LVID;d3 P P P Abbreviations: LVID; left ventricular internal dimension, LVPW; left ventricular posterior wall, IVS; interventricular septum, HR; heart rate, d; diastole, s; systole 193
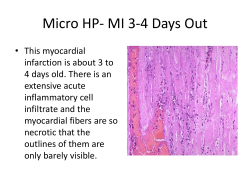









© Copyright 2026