ABC
docz
Explore
Log in
Create new account
Download
Report
health and fitness
disease
cancer
Document 11786
REGULAR LACTATION IN NON-ESTRUS AND ... PREGNANT GOAT FOR YEARS -- A ... Short Communication Shibabrata Pattanayak
EFFECTS OF MILKING FREQUENCY ON MILK PRODUCTION AND
The Carnelian Center
Research Article
Document 11750
Drugs in Milk S
SPLASH!
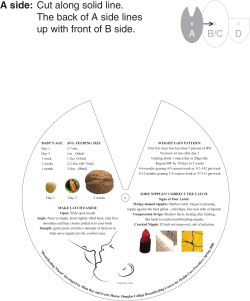
A side: The back of A side lines
FACT SHEET Recombinant Bovine Growth Hormone (rBGH or rBST)
Clinical & Therapeutical study on inactive ovaries hormonal regimes.
© Copyright 2026
About abcdocz
DMCA / GDPR
Report