ABC
docz
Explore
Log in
Create new account
Download
Report
business and industrial
agriculture and forestry
forestry
How to make wildlife conservation more compatible with production forestry:... Kalimantan
Environmentally Sensitive Logging Equipment Qualifications Clear Form Description of Property
SEMEAU 2012 Conference: Forest Management & Water Protection
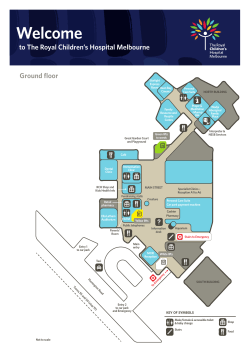
Welcome to The Royal Children’s Hospital Melbourne Ground floor
Research synopsis writing Introduction
Document 51351
EXPERIENCE INVESTMENT MANAGEMENT & REAL ESTATE SERVICES
Forest Fundamentals
June 25, 2014 The Honorable Barack Obama President of the United States
1 Business Plan 2013-2016
© Copyright 2026
About abcdocz
DMCA / GDPR
Report