
Comparison of canine sperm quality under Mestrado Integrado em Medicina Veterinária
Mestrado Integrado em Medicina Veterinária Ciências Veterinárias Comparison of canine sperm quality under different temperature storage Paulo Alexandre Paulos Borges Orientador: Professora Doutora Rita Maria Payan Martins Pinto Carreira Co-orientador: Professor Alain Fontbonne UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO VILA REAL, 2011 Not everything that can be counted counts and not everything that counts can be counted. Albert Einstein ii ABSTRACT Successful gamete cryopreservation is essential to preserve the genetic pool of several species. The canine semen is known to possess a certain resilience to chilling procedures, which has a limited interest for long term gamete preservation; however, when frozen, canine sperm suffers an important decrease in quality. This feature determined the development of different extenders aiming the protection of the cell and to increase the success of canine sperm freezing/thawed procedures. In parallel, development of new techniques allowing a more objective evaluation of putative sperm fertility were also developed. However, those methods do not explain why some stud males are “good freezers” while other are “bad” freezers, when both were selected at start as adequate for freezing. Further, no clear definition exists on the cut-off values for specific seminal parameters to distinguish between a fertile and infertile stud, although the analysis of semen is regularly performed to assess the male fertility. In this study we plan to evaluate cold treatment associated changes on sperm cells by using two different approaches. For that, semen was collected from 8 dogs frequently used as semen donors. Freshly ejaculated samples were assess (Control group) before its use for chilling (Treatment A) or freezing (Treatment B), using routine procedures. For treatment A, three different times were considered: at 1.5h, at 4h and at 24h of chilling (respectively times 1, 2 and 3). Sperm characteristics were assessed by the convention methods, CASA and hypoosmotic test (HOST) for motility and movement parameters, velocity fractions, cell morphology and membrane integrity. Additionally, a molecular approach was tried by the study of heat sock protein 70 (HSP70) immunoexpression in the spermatozoa. HSP70 has protective effects over cells under conditions of thermal or proteotoxic stress. In the study presented here, the more conservative tests confirmed that chilling is not an aggressive procedure for dog semen in comparison to freezing, provided that the samples have a minimum acceptable quality at start. In frozen/thawed samples it was found a decrease in sperm motility and an increase in the percentage of static cells, along with a loss of the sperm membrane integrity. In parallel, a decrease in HSP intensity of immunostaining and a dislocation of the immunoreaction towards the flagellum were observed in the frozen samples (treatment B), while for treatment A no significant changes were found in the pattern of HSP immunoexpression. The use of molecular marker may reprosent an increase value in the study of the mechanisms underlaying the cenine sperm sensitivity to cryopreservation. iii RESUMO A criopreservação de gâmetas é essencial à conservação do pool genético das espécies. O sémen canino é conhecido por ter uma capacidade de resistência à refrigeração aceitável, mas quando congelado, perde potencial fecundante. Esta sensibilidade do sémen canino à congelação incentivou o desenvolvimento de vários diluidores que potencializassem o sucesso da técnica, tendo sido neste processo uma parte integrante o desenvolvimento de métodos relativamente objectivos para avaliação do potencial fecundante do sémen ou dose seminal. No entanto, estes métodos não conseguem explicar os motivos subjacentes à perda de fertilidade observada nem porque alguns machos se apresentam como “bons congeladores” e outros como “maus congeladores” quando todos eles foram seleccionados como adequados para congelar. E apesar de a análise de sémen ser rotineiramente usada para avaliar a fertilidade do macho, não está claramente definido que valores dentro dos parâmetros de sémen distinguem um macho fértil de um infértil. Neste trabalho propusemo-nos a avaliar as alterações associadas ao tratamento térmico pelo frio por dois tipos de metodologia. Assim foram obtidas amostras de sémen de 8 animais dadores regulares de sémen. As amostras foram analisadas a fresco (grupo controlo) e depois processadas de forma rotineira para refrigeração (Tratamento A) e congelação (tratamento B). No grupo de tratamento A foram ainda considerados 3 tempos (tempos 1, 2 e 3, respectivamente às 1.5h, 4h e 24h de refrigeração). O estudo comparativo do potencial fecundante foi realizado por intermédio de métodos convencionais, por CASA e pelo teste hipoosmótico(HOST), para a motilidade e parâmetros de movimento, características de velocidade e integridade funcional da membrana. Esta avaliação foi complementada pelo estudo de imunoexpressão de HSP70 (proteína de shock térmico 70), uma molécula com efeitos protectores sobre a célula em situações de stress térmico e proteotóxico, No trabalho agora apresentado, os testes mais conservadores permitiram confirmar que a refrigeração não é uma técnica muito agressiva para o espermatozóide canino, comparativamente à congelação. Associada a esta última foi observado uma perda na motilidade e na velocidade além de um aumento do nº de células com alteração na função de membrana. Em simultâneo, foi observado um decréscimo na intensidade de marcação para a HSP70, associada a uma deslocação da marcação para a cauda do espermatozóide. A utilização de marcadores moleculares pode revelar-se útil na identificação de mecanismos biológicos subjacentes à sobrevivência do espermatozóide à congelação. iv CONTENTS ABSTRACT .................................................................................................................................. iii RESUMO.................................................................................................................................... IV PART I. ASSESSMENT OF CANINE SEMEN AFTER CHILLING AND FREEZING: A REVIEW .................... 1 1. SEMEN COLLECTION .............................................................................................................. 4 2.1 METHODS OF SEMEN COLLECTION ..................................................................................... 7 2. THE SEMEN ........................................................................................................................... 8 3. THE SEMEN QUALITY ............................................................................................................. 9 3.1 SEMEN ASSESSMENT ....................................................................................................... 10 3.1.1 CLASSICAL METHODOLOGY ....................................................................................... 10 3.1.2 ADVANCED METHODOLOGY ....................................................................................... 15 3.2 SPERM MOTILITY ASSESSMENT ........................................................................................ 16 3.3 HYPO-OSMOTIC SWELLING TEST ...................................................................................... 17 4. HEAT SHOCK ....................................................................................................................... 19 4.1 HEAT SHOCK PROTEINS ................................................................................................... 20 PART II. COMPARISON OF CANINE SPERM QUALITY UNDER DIFFERENT TEMPERATURE STORAGE: EXPERIMENTS .................................................................................................... 21 1. INTRODUCTION AND AIMS ................................................................................................... 22 2. COMMON MATERIAL AND METHODS .................................................................................... 23 2.1 ANIMALS......................................................................................................................... 23 2.2 SAMPLE PREPARATION .................................................................................................... 23 2.3 EXTENDERS..................................................................................................................... 24 2.4 CHILLING PROCEDURE..................................................................................................... 25 2.5 FREEZING/THAWING PROCEDURES................................................................................... 26 3. EXPERIMENTS...................................................................................................................... 29 3.1 EXPERIMENT 1 - EVALUATION OF DOG SEMEN QUALITY AFTER CHILLING AND FREEZING . 29 3.1.1 GOALS ....................................................................................................................... 29 3.1.2 SPECIFIC METHODS .................................................................................................... 29 3.1.3 STATISTICAL ANALYSIS ............................................................................................. 30 3.1.4 RESULTS .................................................................................................................... 30 3.2 EXPERIMENT 2- IDENTIFICATION OF HSP70 CHANGES IN CHILLED AND FROZEN SPERM SAMPLES ................................................................................................................... 39 3.2.1 GOALS ....................................................................................................................... 39 3.2.2 MATERIALS ............................................................................................................... 39 3.2.3 METHODS .................................................................................................................. 39 3.2.4 STATISTICAL ANALYSIS ............................................................................................. 41 3.2.5 RESULTS .................................................................................................................... 41 4.DISCUSSION ......................................................................................................................... 45 5.FINAL CONSIDERATIONS ....................................................................................................... 51 REFERENCES ........................................................................................................................... 52 ANNEXES ................................................................................................................................ 59 ANNEX 1 ................................................................................................................................. 60 ANNEX 2 ................................................................................................................................. 61 v LIST OF FIGURES Figure 1 – Morphological structure of the canine spermatozoon .................................................. 9 Figure 2 – Sperm cell abnormalities (vetmed.lsu.edu) ............................................................... 14 Figure 3 - Semen collection material ......................................................................................... 23 Figure 4 – For preservation, either for chilling or freezing, the collected semen is centrifuged to obtain a pellet with spermatozoa. .................................................... 24 Figure 5 – Sequential procedure of egg yolk addition ................................................................ 25 Figure 6 – From left to right, removal of the supernatant, addition of half of the extender; addition of the second half of the extender ................................................ 25 Figure 7 – Process of storage in the refrigerator ......................................................................... 26 Figure 8 – From left to right, sequential process of supernatant removal, extender addition and storage in the refrigerator ....................................................... 26 Figure 9 - Addition of the second extender ................................................................................ 26 Figure 10 - On the left image, equipment for casing the straws; on the right, procedure of filling up the straws. ............................................................................ 27 Figure 11 - From left to right, procedure of casing the straws..................................................... 27 Figure 12 - From left to right, procedure with the liquid nitrogen ............................................... 28 Figure 13 - Storage of the straws ............................................................................................... 28 Figure 14 - Concentration determination with a spectrophotometer and pH analysis .................. 30 Figure 15 – Immunocytochemical evidence of HSP70 in the spermatozoa head from the control group spermatozoa . ....................................................................... 42 Figure 16 – Immunocytochemical evidence of HSP70 in chilled spermatozoa .......................... 43 Figure 17 - Immunocytochemical staining for HSP70 in frozen/thawed samples ....................... 43 LIST OF GRAPHS Graph 1 – Individual variation of the total motility with treatment ............................................. 32 Graph 2 – Individual variation of the progressive motility with treatment . ................................ 33 Graph 3 - Representation of the sperm total motility (in the left) and progressive motility (on the right) for the group of samples according to the treatment................ 33 Graph 4 – Individual variation of the velocity of the spermatozoa with treatment . ..................... 36 Graph 5 - Individual variation of the hypo-osmotic swelling test (HOST) of the spermatozoa with treatment. ..................................................................................... 38 Graph 6 – Effect of the cryopreservation process on the coiling of spermatozoa......................... 38 Graph 7 – Distribution of HSP 70 intensity in the spermatozoa’s head ....................................... 44 Graph 8 – Distribution of HSP 70 in the spermatozoa’s tail ....................................................... 44 vi LIST OF TABLES Table 1 - The most common causes leading to semen collection in dogs ...................................... 2 Table 2- CASA parameters....................................................................................................... 17 Table 3 – Composition of the extenders used for chilling and freezing/thawing procedures ........ 24 Table 4 – General characterization of the individual semen samples used in this study. .............. 31 Table 5 – Spermatozoa morphology for the freshly ejaculated samples ..................................... 31 Table 6 - Total and progressive motility .................................................................................... 32 Table 7 - Velocity of spermatozoa movement ............................................................................ 35 Table 8 – Hypo-osmotic swelling test ........................................................................................ 37 Table 9 – Immunocytochemical classification of the HSP70 in the spermatozoa ........................ 45 Table on Annex 1 – Total and progressive motility and consequent treatment losses .................. 60 Table on Annex 2 - CASA parameters for sperm cells under cold treatments . ........................... 61 vii List of abbreviations, symbols and units % - percent ® - registered brand AI – artificial insemination ALH - amplitude of lateral head displacement ARTs - artificial reproductive techniques AV – artificial vagina BCF - beat cross frequency CA – California CASA – computerized assisted semen analysis CERCA - Centre d'Étude en Reproduction des Carnivores C-FDA - 6-carboxyfluorescein diacetate DAB - 3,3’-diaminobenzidine tetrahidrocloret DHS – dihydrostreptomycin ENVA - École Nationale Vétérinaire d’Alfort EthD – 1-ethidium homodimer h - hour HOST – hypo osmotic swelling test HSP - heat shock proteins Hz – hertz IL - Illianois IVF – in vitro fertilization KG – kilogram L – liter LIN - linearity of sperm movement ML – mililiter ng – nanogram p – significance level PBS – phosphate-buffered saline PGF2α – prostaglandin F2α PM – progressive motility rpm – rotations per minute s - second STR – straightness SP – seminal plasma TM – total motility UK – United Kingdom USA – United States of America UTAD - University of Trás-os-Montes and Alto Douro VAP – velocity average pathway VCL – curvilinear velocity VSL – velocity straight line WHO – World Health Organization WI – Wisconsin x - average ZBA – zona pellucida binding assay ZP - zona pellucida μm – micrometer μl – microliter viii ACKNOWLEDGEMENTS To both, the University of Trás-os-Montes and Alto Douro (UTAD) and the École Nationale Vétérinaire d’Alfort (ENVA) for all the support given, which allowed the development of this study. To Professor Rita Payan Carreira, my coordinator, I recognize all the dedication applied, the long hours of work, the patience, the friendship, all the wise advices given and the opportunities provided, along with the priceless knowledge she transmitted me throughout my, still short, career. To Doctor Alain Fontbonne, who provided me everything required for the elaboration of my work, along with very important knowledge and opportunities to evolve in the area of small animal reproduction. Within the ENVA and specially the CERCA (Centre d'Étude en Reproduction des Carnivores), I thank Fernando Mir, Emeline Leblond, Cindy Maenhoudt, Natalia Santos and Emmanuel Fontaine for all the attention, knowledge, advices and tender that they offered me during my internship. Also I would like to thank Karine Reynaud and all the laboratory staff that received me very well and helped with everything I needed. To Professor Maria dos Anjos Pires I leave a special acknowledgment, for the special attention and care that she always showed towards me, being available every instant needed and offering all the help and means required. To the Histology and Pathological Anatomy Laboratory of UTAD, Professor Anabela Alves and Professor Maria dos Anjos Pires as its directors along with its technician and staff, Mrs. Lígia Lourenço, Ms. Ana and Mrs. Glória, I thank all the help provided. To my laboratory partners, who were of extreme importance, Inês Santana and Inês Carvalho, I leave a very special thanks for making me embrace the work every day with a smile, for all the teachings, attention, reprimands and specially the friendship provided. ix To all my great friends in Vila Real, Badano, Luis Moreira, Rapper, Verguinhas, Diana, Tatiana, Afilhada, Francisco, Rui, Marta, Ana Margarida, Xami, Zero, João Pires, Magda and Ana Andrade for sharing and giving me so much throughout this so long, though short years. To Professor Wojtek Nizanski who induced and enhanced in me this reproduction thematic and to Natalia Mikolajewska who has been a great support and a very good friend in all the occasions, I thank all the care and preoccupation shown and most importantly the friendship. To Nuno Escudeiro, Vitor Lopes and Ana Lúcia a special thanks for telling me what I need to hear, no matter what, making them true very best friends. The last, but definitely not the least, I would like to thank my family, specially my mother, my father and my little sister, with whom I shared all the tears and joys along my path and who made me the person who I am today most importantly, because without them this would mean nothing. Muito Obrigado! x PART I. ASSESSMENT OF CANINE SEMEN AFTER CHILLING AND FREEZING: A REVIEW 1 FUNDAMENTALS The collection of semen from a male dog may represent an extended procedure during reproductive examination technique. Most frequently, it is used for three major purposes: artificial insemination (AI), semen cryopreservation or diagnostic purposes (Kustritz, 2007). Artificial insemination may be needed in the eventuality of vaginal anomalies of the bitch (such as vaginal septum, vaginal hyperplasia or ptosis, narrowed vagina or vaginal-vestibular stricture), whenever a stud dog is required to produce a seminal dose for breeding at distance or due to other sort of problems, such as behavioral issues between the male and the female (Kutzler, 2005). Canine semen may also be collected in a regular basis for cryopreservation, allowing dog owners to preserve the genetics of their male specimens, thus enabling breeding, even, when the male is no longer able to copulate, if it becomes infertile or if it is physically absent due to the distance or to its death (Johnston et al., 2001; Kutzler, 2005). The most common factors that usually lead to semen collection are described in Table 1. Table 1 - The most common causes leading to semen collection in dogs MOST COMMON CAUSES FOR SEMEN COLLECTION FOR MEDICAL OR OTHER REASONS ASSOCIATED WITH THE REPRODUCTIVE ACTIVITY - Breeding soundness examination - Evaluation of abnormal clinical signs (i.e. prepuce discharge) - Males kept from copulation for several years - History of infertile breedings - Following the treatment of different reproductive tract diseases - AI procedures with fresh or preserved semen - For research - Behavioral issues - Female opposition to the mounting Nowadays, semen cryopreservation and artificial insemination are two of the major biotechnological techniques currently applied in animal breeding (Foote, 2002). Its large-scale introduction in cattle breeding around the 1940’s had the purpose of preventing genital infections transmissible through natural mating (Wilmot, 2007). Semen cryopreservation has also been seen for a long time as a way to improve the breeding of animals of greater farm importance, as a contributor to the conservation of endangered species and also as a way to overcome some aspects of the male infertility (Watson, 2000). Successful preservation of semen is also important to improve the results of the main artificial reproductive techniques (ARTs), such as in vitro fertilization and, most commonly, AI (Luvoni, 2006). Semen cryopreservation eases the transport and exchangeability of genetic 2 material across short or long distances, contributing effectively to widen the restricted gene pool of some species (Johnston et al., 2001; Kutzler, 2005; Kustritz, 2007). Chilling and freezing can also turn into reality the creation of gene banks for particular wild species, which are for the moment maintained under controlled hunting, but may eventually become endangered in the future, as in the case of the Eurasian lynx (Lynx lynx) in Northeastern Europe, or the wild felids in Africa or South America (Swanson, 2006). Cryopreservation of spermatozoa has been well studied in different species, including the dog, having the first pregnancies been successfully achieved with frozen/thawed spermatozoa, using lactose and Tris-(hydroxymethyl)- aminomethane (Tris)-based extenders (Seager, 1969; Andersen, 1972). Canine AI with cryopreserved semen has received increased interest from dog breeders in the past years due to its versatility and to the possibilities that it brings to genetic exchange between distant geographical areas (Thomassen et al., 2009). Furthermore, interest for in vitro technologies for canine species is increasing worldwide and to accomplish the aimed outcomes it is demanded good quality standards for preserved semen (Larsson et al., 2000; Johnston et al., 2001; Kutzler, 2005). The chilling process of dog spermatozoa has been reported to induce less damage in the spermatozoa than freezing (Oettlé, 1986; England et al., 1996). When compared to the observed for the frozen spermatozoa, chilled sperm quality measured by motility, sperm morphology, acrosome status, hypo-osmotic swelling test and longevity, at a temperature of 39ºC, has been reported to be superior, for up to 4.9 days of chilling, despite some deterioration may occur during cold storage (England et al., 1996). Along with motility, the acrosome reaction in dogs seems to be more affected by freezing and thawing than by chilling (Oettle, 1986; Burgess et al., 2001). Recently, few studies have been developed in dogs to determine if the spermatozoa can be chilled and then successfully frozen, having in the end similar post-thawing viability to the semen that follows the normal freezing/thawing procedure (Hermansson, 2006). Cryopreservation of semen, taking into account the process from cooling to thawing, has been proposed to present several negative effects on sperm viability, which could be related to injury of the plasma membrane, associated to changes in lipid phase transition, mechanical stress, efflux of water and high salt solutions and, possibly, by interfering with intracellular ice crystals formation (Woelders, 1997). The type of semen extender and the freezing rates are determining factors on sperm survival during cryopreservation procedures and might explain the variability in the response of canine semen to freezing and thawing, between studies (Peña et al., 2000; Yildiz et al., 2000; 3 Nöthling et al., 2005). The major side effect of the freezing/thawing is a loss of sperm fertilizing ability, mainly associated with reduced motility, promotion of the capacitation and loss of viability (Yildiz et al., 2000). Some of these parameters are associated with sperm membrane damages, which are considered to be the primary cause of freezing induced injuries to canine sperm (Ström Holst, 1998). Furthermore, the same authors also report the existence of significant changes in the elemental composition of post-acrosomal region of the canine spermatozoa after freezing/thawing. In addition, important individual variations were found between stud dogs on what concerns sperm quality after freezing procedures, leading to the common designation of “good” and “bad” freezers (Silva et al., 2003). 1. Semen collection Prior to semen collection a thorough and complete historical review of the dog’s previous health and breeding experiences should be obtained; in addition, information regarding the medication or supplements administered over the previous 6 months (at the minimum) and on the genetic or familiar background could be important (Johnson, 2006). There should also be collection of information about the status of vaccinations, dewormings and heartworm protection history, as well as on the duration of the ownership and accuracy of the history of the animal even before the establishment of the ownership (Freshman, 2002; Ettinger et al., 2010). The anamnesis should be completed with a physical examination, which must consist of a visual, an auscultatory and a palpable examination of the entire animal, with special focus on the parameters or characteristics known to be heritable (CBRA, 1998; Johnson, 2006). This is the moment to talk with the owners and to advise them whether the animal should or should not be bred (Johnson, 2006). This evaluation should be followed by the examination of the reproductive system and it should include the palpation of all the reproductive structures, such as, the scrotum and its contents, the penis and the prepuce and finally the prostate (CBRA, 1998; Simpson et al., 1998). The order of the examination, like the palpation, can and should be postponed till after the collection of the semen, depending on the male’s behavior (Ettinger et al., 2010). The examination of the scrotum and its contents should start with the location of both testes within the scrotum and if that is not the case, the history should confirm a reliable information about the missing testis in order to consider the animal proper for breeding or not (Ettinger et al., 2010). The size and consistency of the testis should be evaluated and registered, along with the position of the tail of the epididymis (to check for example, for a possible torsion of the testicle), as well as the evaluation of the vas deferens, the spermatic cord, the thickness of the scrotal skin and the 4 state of the skin tissues, correlating them with a possible cause of temperature imbalance (CBRA, 1998). The palpation of the prostate should also be included in the physical examination of every male dog, considering its size and shape (Ettinger et al., 2010). Brucella screening tests should also be done in all breeding males (Wanke, 2004). Following the physical examination, it is important to assess the libido, using or not a bitch in heat to facilitate the procedure, but always taking into account that the libido is influenced by several factors, such as the physical conditions of the collection site, the existence of olfactory cues or other specific and unknown reasons related to the bitch used for the effect (Simpson et al., 1998; Johnson, 2006). Many factors influence semen quality, including the animal’s age, the size of the testicles, the degree of sexual arousal, the frequency of ejaculation, the collection procedure and the amount of seminal fluid collected (Johnson, 2006). It is expectable that males may react differently to the surroundings during semen collection and the procedures that precede it (Kutzler, 2005). The collection area should be calm, isolated and interruptions during the procedures should be prevented; the soil should be of secure footing for the animal, using for example a rubber-backed mat that besides providing good footing can also provide olfactory cues to the stud dog (Simpson et al., 1998; Freshman, 2002). Furthermore, in case of a toy dog it could be of benefit to position him on a grooming table (Freshman, 2001; Nelson et al., 2009). The presence of the owner is contradictory, as some dogs can be more comfortable with the owner standing next to them, whilst others can show reluctance to their master’s presence in the room (Purswell et al., 1992; Freshman, 2001). If there are any, the owner should bring the toys or other accessories that the dog associates with the breeding. The collector should avoid any “doctor” paraphernalia, like the white coat or the stethoscope (Freshman, 2002). It is important to remember that the collection of semen should be performed prior to the physical examination, injections, venipunctures or other stressful procedures. As last resort the physical or complementary exams can be even postponed to a different session (Feldman et al., 1996; Johnston et al., 2001). The equipment needed for the procedure should be promptly available and warmed approximately to body temperature (Seager, 1986). Some concerns have been raised on the effect of the usage of latex materials, on the canine spermatozoa motility, for example in the use of artificial vaginas (AV) (Althouse et al., 1991). There are some studies that correlate a decrease in the motility of spermatozoa and the prolonged time of contact with latex, not only with AV but also with gloves; however, this is not an issue when the different techniques are properly chosen and performed (England et al., 1992; Johnston et al., 2001). Another 5 inconvenience of the use of latex collection cones, is the need for a careful cleaning between collections, including the removal of all disinfectant residues, soap and water prior to the next use, taking into account that these are spermicidal (Kutzler, 2005). A sterile non-spermicidal lubricant can be used in minimal quantities in the AV, as so it decreases the friction with the penile mucosa, reducing the risk of injury of the superficial vessels and the penile mucosa (Froman et al., 1983). Previously to the collection procedure, the stud dog should be walked, allowing him to urinate; afterwards, ideally, he should be trotted, to help the cleaning of the urethra from residual urine (Freshman, 2002). The presence of a teaser bitch is usually of benefit, especially if she is in proestrus or estrus and has the same proportions as the stud dog, although a calm bitch can also be used (Freshman, 2002). The typical scent from estrus can either be obtained from frozen swabs or sponges in plastic bags, taken from vaginal secretions (Purswell et al., 1992; Kutzler, 2005) or through the use of a chemical pheromone, methyl p-hydroxybenzoate (Aldrich Chemical, Milwaukee, WI), having them used in the vulvar area and in the tail head of a teaser bitch (Johnston et al., 2001; Kutzler, 2005). It has also been described that administration of prostaglandin F2α (PGF2α; Lutalyse®, Pfizer) in a dosage of 0.1mg/kg, given subcutaneously 15 minutes before the collection, enhances the concentration of the ejaculate, having also a positive effect on the libido. However secondary effects such as salivation, defecation and vomiting can be present due to the use of this drug (Nelson et al., 2009), and the sensitivity of individual males should be tested prior to collection. Allying the additive effects of both a teaser bitch and PGF2α the number of spermatozoa may increase in 300% (Kustritz, 2007). The bitch should be restrained and a muzzle may be used if she seems to be aggressive. The stud dog should be walked behind her, allowing him to sniff and make himself at ease before the collection procedure begins (Schubert et al., 1991). Regarding the collection itself, dog’s prepuce should be vigorously massaged at the region of the bulbus glandis until a partial erection develops, time at which the bulb should be extruded from the prepuce to avoid possible pain for the animal (Freshman, 2001; Kutzler, 2005; Ettinger et al., 2010). If the bulbus glandis becomes too enlarged to be extruded from the preputial opening, removal of the bitch is indicated so that the erection can subside. Once the erect penis is completely extruded, a gentle, firm, circular pressure should be maintained, so that the male dog starts to ejaculate (Ettinger et al., 2010). After the collection, the bitch should be withdrawn from the room and the dog should be monitored till the penis returns to its normal position, inside the prepuce (Freshman, 2002). 6 2.1 Methods of semen collection Semen collection in animals became easier due not only to the development of the AV in 1914 (Gordon, 2004), but also due to the use of the electroejaculator in 1936 (Gunn, 1936). However, nowadays, those methods have been discarded in dogs, in favor of the digital manipulation technique. Besides the digital manipulation of the penis, there are other methods, such as pharmacological ones, that can be used. Nevertheless, the studies that have been done are not very complete, regarding the parameters needed to privilege one method to the others; as an example, an essay has been done consisting on collecting male dogs using pilocarpine hydrochloride dissolved in saline solution, that showed better results than digital manipulation, according to the concentration of spermatozoa, however, no data has been registered relatively to the quality of the collected semen (Juniewicz et al., 1989). Depending on the intended use for the semen, there are some particularities on the way the semen sample should be collected from a dog, taking into account the different fractions that compose the semen. Nevertheless, when there is the need to perform a breeding soundness examination on a dog or if there is suspicion of a reproductive disease, all the three fractions should be collected and evaluated (Kutzler, 2005; Lagishetty et al., 2011). Briefly, if there is the likelihood of risk regarding the fertility, it is important that a complete ejaculation occurs, which can be verified by the analysis of the seminal plasma alkaline phosphatase that should be superior to 10.000 U/L in the combined fractions (Kutzler et al., 2003). For preservation reasons, either semen freezing or chilling, or for fresh semen AI, it may be of benefit to perform two collections, with an interval of 45 to 75 minutes among them. Although the number of spermatozoa is rather low in the second collection comparing with the first, the amount of both collections is in average, 70% more than if the collection is performed only once (England, 1999). The total volume of a dog’s ejaculate may vary from 1.0 mL up to 30.0 mL (England et al., 2010; Ettinger et al., 2010). The canine ejaculate is composed of 3 distinct fractions (Johnston et al., 2001; Kustritz, 2007). The first or pre-spermatic fraction is composed of clear seminal plasma, devoided of sperm cells, originates from the prostatic gland and its main function is to flush the urethra (England et al., 2006; Nelson et al., 2009; Ettinger et al., 2010). The volume of the pre-spermatic fraction usually varies between 0.5 and 2.0 mL (Feldman et al., 1996; Freshman, 2001). The second fraction, also denominated sperm-rich fraction, has a cloudy 7 and opalescent appearance with an opaque consistency, varying in volume between 0.5 and 5.0 mL, depending on the testicular size and on the individual variation; moreover, in its composition there should be no cellular components besides sperm cells (Boucher et al., 1958; England, 1999; Nelson et al., 2009). It originates from the storage in the tail of the epididymis as from the daily sperm output (Johnston, 1989). The dog can take up to 2 minutes to achieve the emission of the sperm-rich fraction (Kutzler, 2005). Ideally there should be a separation between the first and the second fractions, especially for cryopreservation, as it is described that the sperm’s contact with both the first and third fractions may diminish the motility after 2 hours (England et al., 1992). The last portion, the third or prostatic fraction, is usually collected in order to increase the volume of the ejaculate for fresh semen AI and to perform a cytology or a culture (Ettinger et al., 2010). This last fraction can reach up to a total volume of 30 mL, depending on the duration of the pressure maintained around the bulbus glandis (Johnston, 1989). The penile erection can remain for up to 10 minutes and it should be supervised till it wears off, in order to avoid complications (Freshman, 2002; Ettinger et al., 2010). 2. The semen Whenever a semen sample is collected from a dog it should be assessed in order to determine its quality (Kutzler, 2005). At the time of semen collection, information regarding the sexual abstinence should be checked and ideally the stud dog should have 4 to 5 days of sexual rest prior to collection (Freshman, 2002). Nevertheless, it has been reported that more than 10 days of sexual rest can result in an increase of morphologic abnormalities and a decrease in the motility related not only with the ageing of the spermatozoa but also with the increase of debris (Purswell et al., 1992; Johnston et al., 2001). The dog’s ejaculate is composed of cells - spermatozoa (Spz)-, suspended on a fluid - the seminal plasma (SP). Among its constituents, diverse glandular products, proteins and electrolyte were identified (Kustritz, 2007). Several analyses have been performed in order to create a pattern within the normal values of SP mineral contents, specifically zinc, iron, calcium, magnesium and copper (Johnston et al., 2001). Their amounts usually drop till minimum values after the semen collection, due to the metabolism of the spermatozoa and its enzymatic 8 degradation. These parameters can also be influenced by the time interval of the last ejaculation, the sexual arousal of the male and the physiological status of the sexual accessory glands (Kustritz, 2007). Structurally, a spermatozoon can be divided in 3 parts (Figure 1): the head, the midpiece and the tail (England et al., 2010). The sperm head contains a nucleus, which is covered proximally by the acrosome (Cunningham et al., 2007). The midpiece has approximately 1.5 times the length of the spermatozoon’s head (Feldman et al., 2004). A normal dog spermatozoon has approximately 7.0 µm, a midpiece with 1.1 µm and a tail with the length of 5.0 µm divided in its main piece and an end piece (Johnston et al., 2001). Figure 1 – Morphological structure of the canine spermatozoon 3. The semen quality Considering that semen contains live cells, the collector should handle it with special care while executing the procedures, since the interval from the collection till the evaluation should be as brief as possible, in order to prevent cellular modifications or even death of the cells (Feldman et al., 2004). The semen should be maintained between 20ºC to 30ºC (room temperature or water bath) and kept away from direct sunlight (England et al., 2010). Some authors defend that for the analysis of chilled semen, the tubes containing the samples should be kept in a glassed warm water recipient in order to prevent thermal shock during the chilling process, along with temperature variations during the period when the samples stand outside the fridge, for analysis (Iguer-Ouada et al., 2001). Immediately after the end of the collection procedure, the ejaculate should be assessed, since some parameters start to change soon after the collection (Kutzler, 2005). 9 3.1 Semen assessment Each time that a dog is collected with the purpose of insemination, the sample should be assessed in order to determine its quality and to evaluate the reproductive and fertility potential for both the male and the seminal dose (Kutzler, 2005). Usually, available assessment methods vary mainly according to the use of an in-house or a research laboratory on what concerns its profundity or subjectivity (Kutzler, 2005). Although several methods have been proposed for semen evaluation in dogs, the analysis of conception rates should remain as the ultimate test to assess male fertility (Oettle, 1993). The assessment techniques used to determine semen quality may be divided into: - a classical methodology, which include the evaluation of the physical parameters of the collected sample and the assessment of motility, concentration and morphology (Simpson et al., 1998); - a variety of sophisticated assessment techniques, less subjective than the classical, that can be used to assess the motility (like the CASA system) and quality of sperm cells, including the evaluation of the viability, the integrity of the plasma membrane, the capacitating status and the acrosome reaction, among others (Nelson et al., 2009). 3.1.1 Classical methodology The macroscopic parameters in the evaluation of semen are an important component of the classical method and include the volume, the color and the pH (Johnston et al., 2001). The volume, by itself, is not a parameter that defines the quality of the semen, once it depends on the amount of prostatic secretion collected, the age and size of the animal and also the frequency of ejaculations of the dog (Feldman et al., 2004). Nevertheless, it is an essential parameter in order to calculate the total number of spermatozoa in the ejaculate, being of outmost relevance for the semen quality (Simpson et al., 1998). From a laboratorial point of view, the small volume obtained in each ejaculation consists of a great challenge in the semen processing, limiting the number of experiments possible to be done per dose (Farstad, 2009). Usually the appearance of dog’s semen is white, cloudy or opaque and the intensity of its opacity depends on the concentration of spermatozoa (Feldman et al., 2004). The samples that possess a slightly cloudy coloration should be examined under the microscope, to check for the presence of spermatozoa, once that the cloudy color can characterize a sample with no spermatozoa, but can also be associated with a great amount of adipose tissue, bacteria or 10 inflammatory cells (Johnston et al., 2001). The yellow color can indicate the presence of urine, pus or some inflammatory exudates, while the green color normally indicates the presence of pus. The red or brown color can traduce the presence of fresh or haemolyzed blood, respectively (Johnston et al., 2001). The pH should be expected to be within the range of 5.5 to 8.0. Specifically, the normal pH values of the third fraction should vary between 6.0 and 7.4 (Freshman, 2002). As an example, pH measurement may be important regarding the choice of an antimicrobial drug in the eventuality of a prostatitis (Johnston et al., 2001; Freshman, 2002). The microscopic characteristics that should be taken into consideration during the assessment of dog’s semen are included in the protocol used by many andrologic centers and laboratories where light microscopic techniques are routinely used to evaluate the three conventional sperm parameters: concentration, motility and morphology (Johnston, 1991). Semen concentration, along with the volume, is not per se, an indicator of the quality of the semen, as it also depends on the amount of the collected seminal secretion (Johnston et al., 2001; Freshman, 2002). Despite all the technological investment, the method still considered to be the gold standard is also the most traditional one, which uses a counting chamber, such as the Neubauer or the Bürker chambers (Kustritz, 2007). The total number of spermatozoa is achieved multiplying the concentration (millions per mL) by the ejaculate’s volume (mL); the value can vary between 300 million and 2 billion spermatozoa (Ettinger et al., 2010). This wide range of normal values enhances the theory that semen production is directly influenced by the quantity of testicular mass (Johnston et al., 2001; Freshman, 2002; Feldman et al., 2004; Kustritz, 2007). The total and progressive motility can be assessed subjectively on a pre-warmed glass slide and analyzed under a light microscope (Johnston, 1991; Strom et al., 1997). The method should include five different fields of the microscope, in a total of 200 spermatozoa (Kustritz, 2007). The evaluation of the motility can become extremely difficult because of the high concentration of the sample, making it necessary to dilute the semen with a solution that can be the prostatic secretion (natural extender) or a synthetic extender (Feldman et al., 2004; Martinez, 2004). Some saline solutions used as extenders can lead to a decrease on the progressive motility of spermatozoa, due to its pH (Johnston et al., 2001). Other extenders, either due to the presence of viscous substances in their composition (such as egg yolk), the dilution factor or even the temperature at which the analysis is made, may induce a change in the velocity of the spermatozoa, along with the linearity of its movements (Schafer-Somi et al., 2007). The reference value for the progressive motility of dog semen is ≥ 70% (Johnston et al., 2001). The 11 progressive motility of a dog’s semen sample is not affected by the frequency of collections performed (Kustritz, 2007). Nonetheless, the first ejaculate after a long period of abstinence may present a great number of older spermatozoa that have been stored in the epididymis, resulting in a significant decrease in the total spermatozoa with progressive motility in the sample (Feldman et al., 2004). The percentage of morphologically normal spermatozoa is positively correlated with the percentage of spermatozoa with progressive motility (Johnston et al., 2001; Kustritz, 2007), once the motility is considered to be a manifestation of the structural and functional competences of the spermatozoa (Martinez, 2004; Volpe et al., 2009). Although it might not be the best parameter to define the capacity of fertilization of the spermatozoa, motility is an evaluation parameter fairly easy to assess and it can be used as a guideline for the evaluation of fresh and cryopreserved semen (LeFrapper, 2010). Even with all the development around this area and the improvement of more objective techniques for semen evaluation, the progressive motility is still the most used indicator for the spermatic function, once that it is unquestionable that the motility is essential for the progression of the spermatozoa throughout the oviducts (Martinez, 2004). The abnormal motility can be associated with morphologically abnormal spermatozoa and with decreased fertility; however, there can be morphologically abnormal spermatozoa with a normal motility, making it important to evaluate the morphology (Ettinger et al., 2010). The percentage of morphologically normal spermatozoa in a semen sample is considered acceptable over a value of 70% (Feldman et al., 2004). The sperm morphology can be evaluated through the use of different staining techniques, such as eosin/negrosin, Diff Quick or trypan blue (Bangham et al., 1955; Dott et al., 1972; Johnston et al., 2001; Freshman, 2002; Risopatron et al., 2002; WHO, 2010) and it is regarded as an important parameter in the conventional semen analysis (Johnston, 1991; Oettle, 1993). The two most used staining techniques are eosinnegrosin and modified Giemsa (Johnston et al., 2001; Freshman, 2002). The eosin-negrosin as a vital staining has the purpose of emphasizing the borders of the spermatozoa than the cell itself. It also allows, through the use of a contrast phase microscope, the evaluation of the acrosome of live and dead spermatozoa, having the live and acrosome intact spermatozoa a white color with round, regular and defined borders, whilst the live Spz with a reacted acrosome show the acrosomic region in a dark coloration with an unidentifiable apical extremity (Martinez, 2004). This method of coloration allows the differentiation between live and dead spermatozoa, considered as dead ones, the spz that become stained due to the absorption of eosin (staining), 12 once that they are considered to have lesions at the level of the cellular membrane (Kustritz, 2007). It has been described the use of Spermac®. This staining allows a quick staining with unique characteristics, marking the spermatozoon’s nucleus in red, the acrosome, midpiece and tail in green and the equatorial region of the acrosome in pale green (Feldman et al., 2004); this makes this technique a very interesting staining to evaluate the morphology of the acrosome (Martinez, 2004; Monteiro et al., 2009). Apart from the staining technique used, there are always some changes in spermatozoa, resulting from the cytology execution, called artifacts, such as detached heads, coiled or folded tails and folded intermediate pieces, so it can be concluded that the consistency used in the technique is essential for the accuracy and precision of the semen evaluation (Johnston et al., 2001). The use of stainings that are osmotically not similar to the spermatic samples, result in the detachment of the acrosome of some spermatozoa. This osmotic effect is more pronounced in the frozen/thawed than in the fresh semen (Martinez, 2004). Besides the Spz alterations caused by the processing of the semen samples, there are other factors such as testicular traumas, fever or infection of the reproductive tract, that can origin seminal changes, due to the increase of temperature in the testes (Martinez, 2004). However, these changes should only be noted some time after the initial problem, as the spermatogenesis on the dog has a length of 62 days to which the time for epididymal transit should be added (Johnston et al., 2001; Freshman, 2002). Other possible causes for the morphological changes of spermatozoa can be associated with the decrease of LH, a disturbed testosterone secretion or even iatrogenic causes (Martinez, 2004). Healthy dogs that change either of owner or environment can show an increase in the number of abnormal spermatozoa, possibly due to the increase of endogenous corticosteroids (Martinez, 2004). The morphological changes of spermatozoa can be classified according to the cellular location where they occur: head, acrosome, intermediate piece and tail (Figure 2). Furthermore, they can be divided in primary, occurring during spermatogenesis, and secondary if occurring during maturation and for some authors also as tertiary, if during the preparation of the sample (Johnston et al., 2001). Another classification that tries to associate the sperm morphology to the sperm capacity for fertilization, divides the abnormalities of the spermatozoa in minor defects, as those which do not influence the fertility, and major defects, as those which are negatively correlated with the ability to fecundate the oocytes (Johnston et al., 2001). 13 Figure 2– Sperm cell abnormalities (vetmed.lsu.edu) The morphological abnormalities that have been linked to infertility include defects in the connection with the intermediate piece, microcephalic spermatozoa and spermatozoa that possess proximal cytoplasmatic droplets (Feldman et al., 2004). Two main problems that subsist with the light microscopic methods are the subjectivity and the variability (Oettle, 1993; Hewitt et al., 1998). Visual sperm motility assessment is difficult and may be influenced by both the temperature and the evaluator’s skills, leading to high variability among laboratories and observers (Verstegen et al., 2002). On the other hand, the evaluation of the morphology is also problematic due to the fact that it not only depends on the fixation, on the staining technique and on the quality of the microscope, but most importantly it further depends on the observer’s experience and skills (Pena et al., 1999; Pena et al., 1999). The low number of spermatozoa assessed with the conventional techniques is also a factor for inter-laboratory variation (Pena et al., 1998). 14 3.1.2 Advanced methodology Aiming to overcome the principal disadvantages of the current methods of canine semen assessment, other techniques have been proposed, such as fluorescence microscopy, flow cytometry, computerized sperm analysis systems and zona pellucida binding and penetration assays (Gunzel-Apel et al., 1993; Hewitt et al., 1998; Pena et al., 1999; Strom Holst et al., 2000b). One advantage of some of these techniques, such as the use of fluorophore combinations is to evaluate live and dead cells at the same time, either by fluorescent microscopy or by flow cytometry, the last one allowing the evaluation of large numbers of spermatozoa in a short period of time (Rijsselaere et al., 2005). The use of these methods allows also the simultaneous evaluation of several sperm characteristics, relatively to different organelles or sperm domains (Rijsselaere et al., 2005). Wide variations have been described on what concerns the analysis of sperm motility (Chong et al., 1983; Jequier et al., 1983; Mortimer et al., 1986) along with the morphological parameters (Baker et al., 1987; Kruger et al., 1995) of the same ejaculate. The need for higher objectiveness in the methods used, recurring to its standardization for both practical and research purposes, was one of the major concerns in the past two decades (Smith et al., 2001; IguerOuada et al., 2001a). The gathering of these factors opened a window for the development of several semi-computerized (England et al., 1990; Iguer-Ouada et al., 2001b) and computerized measuring devices (Gunzel-Apel et al., 1993; Smith et al., 2001; Iguer-Ouada et al., 2001a; Iguer-Ouada et al., 2001b) for the evaluation of canine sperm motility, morphology and concentration. In order to fertilize an oocyte, a sperm cell should possess the ability to play several functions, including the cascade of reactions that allow the sperm/oocyte interaction at fertilization (Rijsselaere et al., 2005). A great deal of evolution has been gathered around the domains of the assessment of several sperm functions, allowing a more detailed evaluation of dog semen quality (Hewitt et al., 2001). There are some functional assays that are able to examine the interaction between the spermatozoa and the oocyte, thus giving a good estimation of sperm fertilizing capacity, by testing for example the ability of the spermatozoon to bind and penetrate the ZP and fertilize in vitro (i.e. achieved in vitro fertilization, IVF). The ZP binding assay (ZBA) has been used in several species, including the dog (Hay et al., 1997; Strom Holst et al., 2000a; Strom Holst et al., 2000b). In general, the techniques available to assess the semen can be resumed in fluorescent staining, computer-assisted sperm analysis, zona pellucida-binding assay, oocyte penetration assay/in vitro fertilization, and sperm-oviduct interaction (Rijsselaere et al., 2005). 15 3.2 Sperm motility assessment Several commercial computer assisted sperm analysis systems, generally named CASA, such as Strömberg-Mika Cell motion analyzer, CellSoft computer videomicrography; Hobson Sperm Tracker and Hamilton-Thorne, base predominantly on individual spermatozoon assessment and have been validated for use in dogs (Gunzel-Apel et al., 1993; Smith et al., 2001; Iguer-Ouada et al., 2001a; Iguer-Ouada et al., 2001b). These kind of devices allow an accurate and quick calculation of various semen parameters such as the total and progressive motility, the static, slow, medium and rapid moving spermatozoa, the linearity of sperm movement (LIN), the beat cross frequency (BCF), the amplitude of the lateral head displacement (ALH) and quite a few other velocity parameters (Table 2) (Gunzel-Apel et al., 1993; Smith et al., 2001; Iguer-Ouada et al., 2001a; Iguer-Ouada et al., 2001b). There are several studies reporting the existence of high correlations among the computer-calculated motility, progressive motility and concentration, and the conventional light microscopic evaluation (Gunzel-Apel et al., 1993; Iguer-Ouada et al., 2001a). In addition, these systems have proved to be useful when assessing various semen characteristics simultaneously and objectively and are endowed to detect subtle changes in the sperm movement, unnoted with the conventional semen analysis (Gunzel-Apel et al., 1993; Rota et al., 1999; Verstegen et al., 2002). Also, a very important characteristic is that a high number of spermatozoa can be analyzed in a short period of time (Iguer-Ouada et al., 2001a). The main disadvantages pointed out for those devices are the need for high investments and the notorious need for standardization and validation of the software and the system previously to its practical use (Smith et al., 2001; Iguer-Ouada et al., 2001b; Verstegen et al., 2002). Moreover, to avoid a new source of subjectivity among laboratories, associated to the definition of the computer parameters to be applied, the selection of the software and the microscope conditions, the standardization of the technical settings became in focus (Smith et al., 2001; Iguer-Ouada et al., 2001b). In addition, it still needs to be determined which sperm movement characteristics are of best clinical value for the prediction of in vivo fertility in dogs, along with the standardization of the parameters, which differ between users or laboratories and the different commercial systems (Verstegen et al., 2002). 16 Table 2 - CASA parameters (modified from Iguer-ouada et al., 2001a; Verstegen et al., 2002; Rijsselaere et al., 2005) Parameter Definition TM Total motility PM Progressive motility VAP Velocity average pathway VSL Description Unit Percentage of spermatozoa with movement % Percentage of spermatozoa that progress; STR > x (straightness threshold cut-off to determine the progressive spermatozoa) % Average velocity of the smoothed cell’s pathway μm/s Velocity straight line Average velocity measured in a straight line from the beginning to the end of one track μm/s VCL Curvilinear velocity Average velocity measured over the actual point-to-point track followed by the cell μm/s ALH Amplitude of lateral head BCF Lateral amplitude of the head displacement μm Beat cross frequency Frequency at which the sperm cell’s head crosses the sperm cell’s average pathway Hz STR Straightness Average value of the ratio VSL/VAP in percentage (estimates the proximity of the cell’s pathway to a straight line with 100% corresponding to the optimal straightness % LIN Linearity Average value of the ratio VSL/VCL in percentage (estimates the proximity of the cell’s track to a straight line) % The CASA is basically a computer analyzer for sperm motility and morphometry. Its use has become more popular, but due to its high cost it is still not available to the majority of the veterinarians (Verstegen et al., 2002; Kustritz, 2007). With a contrast microscope integrated, this equipment allows a quick and objective evaluation of many parameters, some of them described in Table 2. As this equipment allows to identify even very small variations in the spermatozoa motility, that would, otherwise be impossible to detect (Rijsselaere et al., 2005), it allows the classification of small populations of spermatozoa in a sample that may react differently when submitted to processes, such as cryopreservation (Kustritz, 2007). 3.3 Hypo-osmotic swelling test The morphological and functional integrity of the sperm membrane has been intensely studied, given its importance as a cellular barrier and in the cell to cell interaction (RodríguezMartínez, 2000). The integrity of the plasma membrane is crucial for the fertilizing capacity of spermatozoa. As described to the morphology, in the classical methods, the membrane of dog’s 17 spermatozoa was routinely assessed by means of light microscopic stainings, such as eosin/negrosin (Bangham et al., 1955; Dott et al., 1972) or trypan blue (Risopatron et al., 2002). An indirect method to evaluate the membrane integrity is by exposing spermatozoa to hypoosmotic conditions, for example through the hypo-osmotic swelling test (Kumi-Diaka, 1993), once that the number of swollen spermatozoa was shown to be inversely proportional to the number of membrane damaged spermatozoa (England et al., 1993). This test is considered, by many laboratories, as an appropriate method to evaluate the integrity of the membrane and due to its simplicity, it can be introduced into the routine semen analysis (Pinto et al., 2008). This technique consists of mixing the semen sample in a hypoosmotic solution and incubating it at 37ºC, for 45 to 60 minutes (Rodríguez-Gil et al., 1994). When submitted to a hypo-osmotic solution, a viable spermatozoa possessing an intact, active and functional plasmatic membrane, will react, allowing the fluid to enter the cell, causing the swelling and coiling of the tales of spermatozoa (Kustritz, 2007). It has been proved that no significant differences exist between incubation times of 1 or 60 minutes, and also that this test is easily performed by any veterinarian (Pinto et al., 2008). The solution used in this test should be enough hypo-osmotic to cause the swelling, without causing lysis to the cell (Bencharif et al., 2008). The hypo-osmotic solutions described for this test include sodium citrate, fructose in distilled water and a solution of sacarose (Kustritz, 2007). The swelling effect is more easily identifiable on the tails of the spermatozoa for two main reasons: the membrane of the tail is more flexible and is less adherent to the internal structures comparing to the membrane of the head, and at the spermatozoon’s head the intracellular fluid compartment is rather small making it difficult to observe the variation in the volume when the water enters the cell (Jeyendran et al., 1984). Previously to the realization of the hypo-osmotic swelling test, it is of high relevance that the percentage of spermatozoa with tail abnormalities is measured, in order to subtract this value to the counting made after incubation, obtaining the true value of spermatozoa with an intact plasmatic membrane (Kustritz, 2007). This test shows a good correlation among the spermatozoa that suffer swelling and the percentage that were able to fertilize the oocytes in vitro. In the same study, the correlations obtained between this parameter, the percentage of progressive motility, the percentage of morphologically normal spermatozoa and the percentage of live spermatozoa (negative with vital staining) was not so relevant (Jeyendran et al., 1984). There is no relation described between the 18 hypo-osmotic swelling test results and the fertilization capacity of dog spermatozoa (Martinez, 2004), however there is a correlation between these results and the motility and viability for both fresh and frozen dog semen (Pinto et al., 2008). 4. Heat shock A reaction to an induced heat shock is inherent to the procedure of semen cryopreservation, which is accompanied by the irreversible loss of viability when the spermatozoa rapidly reach neutral temperatures. This co-exists with a decrease in sperm motility (Quinn et al., 1966), a reduction in the energetic ability and an imbalance of the membrane’s permeability (Watson, 1981). Thus, the refrigeration of the semen during the freezing process should be done with extreme precaution, in a slow procedure, even if it does not prevent all the damage to the membrane (Watson, 2000). It has been reported that the spermatozoa become susceptible to the heat shock in the proximal region of the body of the epididymis, during the epididymal maturation process; more precisely it occurs when the cytoplasmatic droplet moves towards the distal portion of the midpiece. It has been concluded that acquisition of motility and heat shock susceptibility may occur at the same time (White, 1993). The freezing-associated heat shock and the irreversible lesions that it causes, seem to result from the changes in the lipid organization in the sperm cell membrane (Drobnis et al., 1993). The heat shock has different impact according to the species, being the sperm cells of both swine and bull the most susceptible. There are relevant changes in the ion exchanges, with Ca2+ increase and loss of K+ and Mg2+. On the other hand, the sperm of humans, dog and rabbit seems not to show so intense changes (Quinn et al., 1966). Nevertheless, freezing/thawing processes in canine semen are usually referred as being detrimental to the sperm fertility, mainly due to reduced longevity (England et al., 1993; Bessa, 2005) and to an increase in the proportion of capacitated cells after thawing (Rota et al., 1999; Bessa, 2005). These issues were at the origin of intense scientific work aiming to identify the most adequate extender for dog semen. 19 4.1 Heat Shock Proteins Heat shock as other stressing factors stimulate every cell to synthesize a small group of protective proteins, which are named heat shock proteins (HSPs) (Schlesinger et al., 1982; Craig, 1985; Schlesinger, 1986). HSPs are classified into several families, named according to their approximate molecular weight (Sonna et al., 2002). Among several stress stimuli, elevated temperature or heat stress have been described as inducers of the HSPs production in certain tissues (Nollen et al., 1999). The concentration of stress proteins increase, responding to stress factors, therefore acting as cell protectors and facilitating cell survival, proportioning renaturation of partially denaturated proteins (Parcellier et al., 2003). There have been important and relevant studies about the heat shock proteins throughout the last years. These studies have been more incisive in a special set of evolutionary conserved heat inducible and related proteins of molecular weight of around 70 kDa (Lawson et al., 1984; Craig, 1985; Schlesinger, 1986). HSPs, and specially HSP70, under physiological conditions act as molecular chaperones as well as assisting protein folding, transport and degradation; on the other hand, subjected to stress conditions, they prevent aggregation and promote refolding after denaturation, protecting cells of stress or other hazardous conditions (Georgopoulos et al., 1993). Induction of heat shock response is related to the physiological temperature range (Liu et al., 1994) and can occur either when the temperature increases upwards a threshold or when it decreases below a specific value. Responding to heat stress, heat shock genes are stimulated and new molecules of HSP are produced within the cells, in an attempt to prevent or to correct denaturing of cell proteins. Thus, induction of the heat shock response should be read as a manifestation of temperature-induced cell damage (Liu et al., 1994). One important implication of this finding in sperm post-thawing fertility is that events involved in the expression of the heat shock response require active cell metabolism, such as ATP production or macromolecule synthesis, which are of limited capacity in sperm cells due to their highly specialization. 20 PART II. COMPARISON OF CANINE SPERM QUALITY UNDER DIFFERENT TEMPERATURE STORAGE: EXPERIMENTS 21 1. Introduction and aims The specific thematic for this work was chosen accordingly to the work developed throughout the 3.5 months of practical training in the École Nationale Vétérinaire d’Alfort, in the area of small animal reproduction, CERCA (Centre d'Études en Reproduction des Carnivores), which is a highly regarded centre for reproduction and specifically cryopreservation. In the process of cryopreservation, the sperm cells, in particular dog sperm cells are highly susceptible and suffer substantial alterations, especially on what concerns sperm motility. As proposed aims for this study, we ought to: 1 – Evaluate the sperm quality upon chilling and freezing/thawing procedures and to compare it to the freshly ejaculated sperm. 2 – Characterize the pattern of immunolocalization of HSP70 in the freshly ejaculated canine spermatozoa and to study putative changes after being chilled and frozen/thawed. We further will attempt to study eventual association between data gathered in those two studies, and to ascertain possible influences of the temperature stress induced by the two cold treatments on sperm quality. To achieve the proposed objectives we performed two different studies where the same samples were used. For the first experiment, freshly ejaculate samples were obtained from 8 dogs. Those samples were processed in straws as for routine seminal doses and submitted to chilling for 24h and to freezing in liquid nitrogen with posterior thawing. The quality of each sample was assessed by the use of both classical and advanced methods, for each temperaturechallenge tests, compared to controls (fresh ejaculates) and differences among treatments and individual dogs were analyzed. In this study, the following parameters were evaluated: concentration, pH, volume, color, morphology and HOST, as well as CASA characteristics (total and progressive motility, VAP, VSL, VCL, ALH, BCF, STR, LIN). For the second experiment, cytological specimens were prepared from each sample, fixed at 95% ethanol and processed for immunocytochemistry. The characterization of the normal expression pattern of HSP70 was studied in the control samples (fresh ejaculates). The intensity of immunostaining as well as the sub-cellular location of the protein was recorded. Comparison between controls and the cold-treated groups (chilling and freezing/thawing) was evaluated on what concerns the intensity of immunoreactions and any possible changes in the sub-cellular location of HSP70. With this approach we ought to test the possible involvement of HSP70 in some treatment-associated changes found in more conventional sperm assessment. 22 2. Common material and methods 2.1 Animals In this work, fresh ejaculates were obtained by digital manipulation, according to the method described by Linde-Forsberg (Linde-Forsberg, 1991), from 8 dogs, with ages ranging between 3 to 4 years, that either belonged to the École Nationale Vétérinaire d’Alfort, Paris or were received in CERCA (Centre d'Études en Reproduction des Carnivores) for consultation. There was no specificity neither according to the breed chosen, nor the age of the dogs, besides that all of them were considered to be healthy and reproductively within the accepted fertility parameters, since a reproductive exam and a prostatic ultrasound were previously performed. The collected sample included the sperm-rich (second) fraction and a small amount of the third fraction Figure 3. Figure 3 - Semen collection material 2.2 Sample preparation For chilling procedures, after dilution the samples were stored in test tubes in the refrigerator for up to 24h. For frozen samples, 0.50 mL straws were used and stored in liquid nitrogen for a period of 2 days. From fresh ejaculates, and after semen evaluation, the straws were prepared in order to obtain a concentration of 150 x 106 spermatozoa in each straw. The following formula was applied: 23 Fresh samples were centrifuged for 10 minutes at 1.500 rpm, as displayed in figure 4. The exceeding fluid was withdrawn and only the sperm pellet was kept. The pellet was then resuspended in the adequate extender, according to the temperature treatment to be followed: chilling or freezing. Figure 4 – For preservation, either for chilling or freezing, the collected semen is centrifuged to obtain a pellet with spermatozoa. 2.3 Extenders The extenders used in the study, for both chilling and freezing/thawing of the semen, correspond to the method of Uppsala, (Linde-Forsberg et al., 1999). The extenders were prepared with Tris, citrate, glucose, antibiotics, glycerol, Equex and distilled water (Table 3) and were frozen. Table 3 – Composition of the extenders used for chilling and freezing/thawing procedures Components Chilling extender Freezing extender 1 Freezing extender 2 Thawing extender TRIS 6.025 g 3.025 g 3.025 g 3.025 g Citrate 3.4 g 1.7 g 1.7 g 1.7 g Glucose - 1.25 g 1.25 g 1.25 g Fructose 2.5 g - - - Peniciline 200 000 UI 0.06 g 0.06 g 0.06 g DHS 0.2 g 0.1 g 0.1 g 0.1 g Distilled water Till 200 mL 77 mL 72 mL Till 100 mL Glicerol 12 mL 3 mL 7 mL - Equex - - 1 mL - Egg yolk 20 mL 20 mL 20 mL - 24 The preparation was made in advance, by mixing the powder components in water and, for the extender 2, the final adding (without mixing) of the glycerol or the Equex. It was filled up to 100 mL or 200mL of water, according to the extender, and it was strongly mixed. It was stored in plastic tubes in the amount of 4 mL each and frozen to future use. At the time of use 1 mL of egg yolk was added to both extenders (Figure 5), having a total of 5 mL of each extender to use for the chilling or the freezing procedure. Figure 1 – Sequential procedure of egg yolk addition 2.4 Chilling procedure After the withdrawn of the supernatant with a Pasteur pipette, one half of the calculated chilling extender was added to the sperm pellet. After waiting one minute, the second half of the extender was added, with the proper care of not jetting it harshly, as shown on figure 6. It was stored in the refrigerator or chilling chamber, as displayed in figure 7, for 24 hours at 4ºC, having been collected small samples to analyze at the time of 1.5, 4 and 24 hours. Figure 6 – From left to right, removal of the supernatant, addition of half of the extender; addition of the second half of the extender 25 Figure 7 – Process of storage in the refrigerator 2.5 Freezing/thawing procedures After the supernatant removal, with a Pasteur pipette, the calculated freezing extender 1 was added to the tube with the sperm pellet, and then stored in the refrigerator or the chilling chamber for 1.5 hours at 4ºC (Figure 8). In a separate container, the second extender was also placed in the refrigerator, to chill and equilibrate the temperature with the pre-diluted sperm sample. Then, the second extender was carefully added, avoiding jetting it harshly (Figure 9). Figure 2 – From left to right, sequential process of supernatant removal, extender addition and storage in the refrigerator Figure 3 - Addition of the second extender Immediately after mixing, the semen was shed in a leaned metallic container, in order to case the semen in straws. The end of each straw with no cotton was submerged and the semen 26 was aspirated, with a pipette or another aspiration method, as shown in Figure 10. A small air chamber was left inside each straw, to prevent its explosion during freezing. To seal the straw, the end devoided of cotton was inserted in a specific powder for several times and then submerged into water, to become solid, as displayed in Figure 11. Figure 4 - On the left image, equipment for casing the straws; on the right, procedure of filling up the straws. Figure 5 - From left to right, procedure of casing the straws The straw was then placed under nitrogen vapors for 20 minutes, and therefore completely submerged in the liquid nitrogen, as seen in Figure 12. The straws for this study were finally stored in a specific nitrogen container for 2 days, as shown in Figure 13. 27 Figure 6 - From left to right, procedure with the liquid nitrogen Figure 7 - Storage of the straws For thawing, the straws were submerged directly in a water bath at 37ºC, for one minute, and dried up afterwards. Its powder end was cut off, while the remaining of the straw was kept inside a plastic tube already containing 0.5 mL of pre-warmed PBS (phosphate-buffered saline solution). The cotton end was then also cut off to allow the seminal doses to flow. 28 3. Experiments 3.1 Experiment 1 - Evaluation of dog semen quality after chilling and freezing 3.1.1 Goals In this study, we ought to evaluate the differences in sperm quality following two different temperature challenges (chilling vs. freezing/thawing), using fresh ejaculated semen as controls (Time zero). For this experiment, we considered 3 different times during chilling treatment (at 1.5h, 4h and 24h, respectively treatment A, times 1, 2 and 3) and the freezing treatment (treatment B). 3.1.2 Specific methods Fresh ejaculates were assessed by classical methodology, the CASA technology and the hypo-osmotic swelling test. Temperature-treated samples were subjected to both CASA device analysis and hypo-osmotic swelling test. The macroscopic parameters evaluated included the semen volume, color and pH. Also, the morphology of the spermatozoa was evaluated in a cytology stained with eosin-nigrosin. The abnormal forms found were classified according to Johnston (Johnston et al., 2001). Within the microscopic parameters the concentration of the seminal sample was determined with a spectrophotometer, whilst the morphology was assessed in eosin-negrosin stained slides (Figure 14). In order to maintain the coherence, massal and individual motility parameters were assessed by CASA, as for the treatment samples. All the semen samples were left for 2-3 minutes to warm up, in a warm bath at 37ºC. A volume of 3μl was placed, with the help of a micropipette, in a pre-heated Leja® standard count 4 chambers slide of 20 micron and the assessment with the CASA device was initiated 1 minute after placing the Leja® slide in the warmed plate of the microscope. The parameters taken into consideration from the CASA analysis were the total and progressive motility, the velocity of the Spz movement (rapid, medium, slow, static) and the different characteristics of Spz agility, VAP, VSL, VCL, ALH, STR and LIN. 29 Figure 8 - Concentration determination with a spectrophotometer and pH analysis 3.1.3 Statistical analysis Data gathered in the two experiments were organized in Microsoft Excel . Statistical comparisons were performed by using the IBM SPSS Statistics Base 19.0 statistical software for Windows®. Statistical analysis of the effects of treatment on motility, velocity and HOST were performed using the ANOVA, with Bonferroni adjustment. A p value ≤ 0.05 was regarded as statistically significant. 3.1.4 Results Semen samples collected from our 8 dogs showed variable individual semen concentrations (Table 4). DPAR’s semen was the less concentrated, with a 70x106 Spz/mL, in contrast to the one from ELWOOD, which was the most concentrated, with a value of 657x106 Spz/mL (Table 4). The collected seminal volume, by gathering both the 2 nd and part of the 3rd fraction, varied between 2.5 and 5 mL (Table 4). The samples showed a pH within the normal range for the canine semen (between 6 and 6.5, as shown in Table 4) and remained unchanged throughout the cryopreservation process. The color of all the semen samples collected was opaque white (Table 4), aiding in this way to prove the good reproductive status of the animal and not evidencing a bad method of semen collection. Considering the morphology of the spermatozoa, all the samples showed acceptable and similar percentages of normal spermatozoa, with DPAR´sample being the one with the lowest percentage of abnormal sperm forms. DINGO´s sample was the exception as it presented a higher amount of abnormal sperm forms when compared to our other samples and even to reference values. In this individual, it was particularly notorious the high percentage of tail defects. 30 Table 1 – General characterization of the individual semen samples used in this study. Animals Concentration (x106 Spz/mL) Volume (mL) pH Color DPAR 70 4 6 Opac white DLIT 261 2,5 6 Opac white ELTON 421 3 6.5 Opac white ELIOS 200 5 6.5 Opac white TECH 186 4 6.5 Opac white DINGO 114 4 6.5 Opac white UROCH 309 3 6 Opac white ELWOOD 657 3 6 Opac white Table 2 – Spermatozoa morphology for the freshly ejaculated samples (control group) Animal DPAR DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD % Normal % Abnormal Abnormal heads (n) Intermediate pieces (n) Abnormal tails (n) Droplets (n) Detached heads (n) 98,5 89,5 87 90,5 91 77,5 93 93 1,5 10,5 13 9,5 9 22,5 7 7 0,5 1,5 3 5 0,5 3 2 3 0 5 4,5 2 1,5 2 2 1 0 3 4 2 4 16 2 3 0,5 0 0,5 0,5 1 0 0 0 0,5 1 1 0 2 1,5 1 0 The results of total and progressive motility of all the 8 dogs can be observed in Table 6 and more clearly in Graphs 1 to 3. Regarding the evaluation of spermatozoa motility in the CASA system, in the control group ELTON and DINGO´s samples showed the lowest total motility values and consequently also the lowest results for the progressive motility. For Treatment A (chilling), no overall significant variations were observed, having the majority of the animals better total motility for time 3 (24h chilled) than time 2 (4h). The one exception was DLIT, which showed a slight decrease in the total motility and lower values for progressive motility at time 1 (1,5h chilled) (Graphs 1 to 3; Table 6).In treatment B group a significant decrease in the motility parameters were observed in all the samples of the group. Again, the most deleterious effects of freezing/thawing over sperm motility were found for DINGO´s (Graphs 1 to 3), with a total and progressive motility of the frozen samples reduced to 16 % and 12% respectively (Table 6). 31 Table 3 - Total and progressive motility Total motility (%) Samples Controls DPAR Treatment A Treatment B Time 1 Time 2 Time 3 89 86 85 84 71 DLIT 85 66 67 69 53 ELTON 68 86 72 81 53 ELIOS 80 87 80 88 55 TECH 89 76 81 85 50 DINGO 61 82 71 81 16 UROCH 88 81 87 87 60 ELWOOD 73 81 77 89 61 Progressive motility (%) DPAR 61 49 52 59 39 DLIT 61 39 40 44 34 ELTON 49 49 53 53 43 ELIOS 54 49 52 50 33 TECH 55 53 52 47 31 DINGO 32 58 50 53 12 UROCH 52 52 54 47 32 ELWOOD 50 51 53 54 43 30 20 10 0 DPAR DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD -10 -20 -30 -40 -50 Chilling T1 Chilling T2 Chilling T3 Freezing Graph 1 – Individual variation of the total motility with treatment, compared to control samples (Time 0). The units in Y-axis correspond to the unitary differences between the total motility (%) between the temperature treated samples (Chilling time 1, 2 and 3, and freezing/thawed) and the controls. 32 30 20 10 0 DPAR DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD -10 -20 -30 Chilling T1 Chilling T2 Chilling T3 Freezing Graph 2– Individual variation of the progressive motility with treatment, compared to control samples (Time 0). The units in Y-axis correspond to the unitary differences between the progressive motility (%) between the temperature treated samples (Chilling time 1, 2 and 3, and freezing/thawed) and the controls. Graph 3- Representation of the sperm total motility (in the left) and progressive motility (on the right) for the group of samples according to the treatment. Motility values were obtained from the CASA analysis. The central bar means the median value, the lower line stands for the lowest value, while the upper one, its opposite. Outside values correspond to samples considered to be “outsiders” to the core population. 33 Cold treatment was found to significantly influence the total motility as well as the progressive motility (p<0,0001). However, those differences were found only for freezing/thawing group in comparison to chilling (treatment A) and to controls. No differences were found between the controls and treatment A group concerning the total motility or the progressive motility. The velocity of movement of spermatozoa was also evaluated and scored as rapid, moderate, slow and static, by using the CASA system (Table 7). The results from this evaluation reinforce the information provided by the motility assessment. On what concerns this parameter, both ELTON and DINGO samples showed lower percentage of rapid spermatozoa, 61 and 50% respectively, regarding the control group. Regarding the treatment A group, at time 1 DLIT shows the most important decrease in the rapid spermatozoa, from 81% to 55%, while both ELTON and DINGO show very acceptable values, an increase in these values being noted (78 and 76%). According to the treatment B, the animal that shows a higher decrease is TECH with a decrease of 42% of rapid spermatozoa considering the control group (Table 7). For the analyzed CASA parameters (VAP, VSL, VCL, ALH, STR and LIN), only averages were taken into consideration (Table 8). On what concerns the velocity of average pathway (VAP), small changes were observed in treatment A group, with half of the animals presenting a decrease (DPAR, DLIT, ELTON and UROCH), which was more pronounced at Times 2 and 3, whilst the other half showed an increase (ELIOS, TECH, DINGO and ELWOOD). Regarding the treatment B, all the samples presented a decrease in VAP value, with the exception of TECH and DINGO´ samples. A similar situation was found for the velocity of straight line (VSL), where for both treatment A and B groups the samples from the abovementioned animals showed similar fluctuations. Attending the curvilinear velocity (VCL) for treatment A, a decrease in values was found for Time 1 in DLIT, TECH and UROCH´ samples, which were also registered towards Time 2 for DLIT, ELTON and UROCH. For Time 3, all the samples showed increased values in comparison to controls. In contrast, for treatment B all the samples used showed a decrease in VCL values (Table 8). 34 Table 4 - Velocity of spermatozoa movement Rapid (%) Samples Controls DPAR Treatment A Treatment B Time 1 Time 2 Time 3 84 78 79 79 60 DLIT 81 55 57 60 44 ELTON 61 78 66 75 49 ELIOS 75 82 76 82 47 TECH 83 71 76 79 41 DINGO 50 76 65 76 13 UROCH 83 74 81 82 47 ELWOOD 67 75 71 84 53 Medium (%) DPAR 4 8 6 5 11 DLIT 4 10 10 8 9 ELTON 7 8 6 6 3 ELIOS 5 6 4 6 8 TECH 5 5 5 6 9 DINGO 11 5 6 5 3 UROCH 5 7 6 5 13 ELWOOD 6 7 7 5 8 Slow (%) DPAR 12 13 14 14 28 DLIT 14 32 31 24 41 ELTON 31 14 25 18 36 ELIOS 20 12 19 12 40 TECH 11 23 18 15 46 DINGO 30 17 26 16 26 UROCH 12 18 12 12 38 ELWOOD 27 17 20 11 26 Static (%) DPAR 0 1 1 2 1 DLIT 1 3 2 8 6 ELTON 1 0 3 1 12 ELIOS 0 0 1 0 5 TECH 1 1 1 0 4 DINGO 9 2 3 3 58 UROCH 0 1 1 1 2 ELWOOD 0 1 2 0 13 35 30 Medium velocity fraction Rapid fraction 20 10 0 -10 -20 -30 -40 -50 Chilling T1 60 Chilling T2 Chilling T3 Freezing Static fraction Slow fraction 50 40 30 20 10 0 -10 -20 -30 Chilling T1 Chilling T2 Chilling T3 Freezing Graph 4– Individual variation of the velocity of the spermatozoa with treatment, compared to control samples (Time 0). Upper: for rapid and medium velocity fractions; bottom: for the slow and static fractions. The units in Y-axis correspond to the unitary differences between the total motility (%) between the temperature treated samples (Chilling time 1, 2 and 3, and freezing/thawed) and the controls. When evaluating the lateral amplitude of the head (ALH), in treatment A group, there was found a decrease in TECH, DINGO and UROCH’s values at Time 1, with only TECH and DINGO remaining with lower values at Time 2. At Time 3 an increase of ALH values was found for most samples used, with exception of ELWOOD´s which registered a pronounced decrease for 36 this parameter. In the treatment B group, DLIT and ELIOS´ samples failed to evidence a decrease in ALH values. Concerning the straightness (STR) movements of the spermatozoa, at the initial Time for treatment A group samples TECH, DINGO and UROCH showed an increase toward the controls, with only Dingo´s sample maintaining such increase by Time 3. For treatment B group, both DPAR and ELIOS showed lower STR-values in comparison with the controls. For the spermatozoa’s linearity (LIN), in the treatment A, an increase in the values was found for samples from TECH, DINGO and UROCH at Time 1, and for TECH and DINGO in Time 2, which were maintained in Time 3. For treatment B, only samples from ELTON, TECH and DINGO showed an increase in LIN values (Table 8). Regarding the hypo-osmotic swelling test (HOST) the largest variation in terms of coiled spermatozoa were noted in DINGO comparing the frozen and fresh semen, while for the chilling process, a decrease was also noted though in an equilibrate way among all the animals (Table 9). Throughout the chilling process a slight decrease was noted in the animals, apart from DINGO, which presented a small increase in coiled spermatozoa at 24h (Graph 5). No statistical differences were found between the controls and the group for treatment A (p>0.05), no matter the time considered, however both the controls and treatment A group statistically differed from the treatment B group (p<0.0001). Table 5 – Hypo-osmotic swelling test DPAR DLIT ELTON ELIOS TECH DINGO UROCH Fresh 90.9 94 95 97 92 88 95 ELWOOD D 96 Chilled 1.5h 96 84 89.1 86 93 93 94 93 Chilled 4h 96 85 89 98 92 91 93 96 Chilled 24h 96 79 89 86 89.1 97 91 93 Frozen 74 50 57 49 45 31 56 49 Fresh 9.1 6 5 3 8 12 5 4 Chilled 1.5h 4 16 10.9 14 7 7 6 7 Chilled 4h 4 15 11 2 8 9 7 4 Chilled 24h 4 21 11 14 11 3 9 7 Frozen 26 50 43 51 55 69 44 51 Hiposmotic Test HOST (coiled) HOST (not coiled) 37 Graph 5- Individual variation of the hypo-osmotic swelling test (HOST) of the spermatozoa with treatment, compared to control samples (Time 0). The units in Y-axis correspond to the unitary differences between the total motility (%) between the temperature treated samples (Chilling time 1, 2 and 3, and freezing/thawed) and the controls. Hyposmotic swelling test 100 75 Not coiled 50 Coiled DPAR C DLIT C ELTON C ELIOS C TECH C DINGO C UROCH C ELWOOD C DPAR R 24 DLIT R 24 ELTON R 24 ELIOS R 24 TECH R 24 DINGO R 24 UROCH R 24 ELWOOD R 24 4 4 4 4 4 4 4 4 DPAR R DLIT R ELTON R ELIOS R TECH R DINGO R UROCH R ELWOOD R DPAR F DLIT F ELTON F ELIOS F TECH F DINGO F UROCH F ELWOOD F 0 DPAR R 1,5 DLIT R 1,5 ELTON R 1,5 ELIOS R 1,5 TECH R 1,5 DINGO R 1,5 UROCH R 1,5 ELWOOD R… 25 Graph 6 – Effect of the cryopreservation process on the coiling of spermatozoa. 38 3.2 Experiment 2- Identification of HSP70 changes in chilled and frozen sperm samples 3.2.1 Goals In this study, we proposed ourselves to evaluate the presence of the HSP70 in the canine spermatozoa. Another purpose of this project was to observe the distribution and relocation of the HSP70 on the canine sperm, under the influence of two different cryopreservation processes, such as chilling and freezing, treatment A and B, respectively. The final aim was to compare the distribution of the HSP70 and the quality of the semen, after the two cryopreservation procedures. 3.2.2 Materials For the purpose of immunocytochemical analysis, 8 semen samples were collected and processed, submitting them to a smear procedure in silane coated slides (3- Aminopropyltriethoxysilane, Sigma ®), according to the conventional methodology and fixated in alcohol at 95%. For each cryopreservation procedure 10 slides of the different samples were prepared. In this study, HPS70 immunoreaction was study in samples from treatment A, at 24h of chilling (time 3) and from treatment B, using the fresh semen samples as controls for the normal intensity and pattern of immunoreaction. The expression of the proteins in the study was evaluated by the indirect method of the Streptavidin biotin-peroxidase, using the solution 3,3’-diaminobenzidine tetrahidrocloret (DAB) as the cromogenic agent. The immunocytochemical expression of the antibodies was obtained through the indirect method with the use of the complex streptavidin-biotin peroxidase, using the kit Ultravision Detection System, Labvision Corporation®, Fremont, CA, USA. 3.2.3 Methods 3.2.3.1 Immunocytochemical study In order to make the technique as accurate as possible, preliminary studies were performed, so that the dilution of the antibodies could be determined, the method of the antigenic recuperation chosen and the different times of the incubation period analyzed, optimizing 39 through these steps, the final results. These trials allow us to identify the most suitable antigen retrieval method and primary antibody dilution. The silane coated slides were hydrated through a series of alcohols with decreasing concentrations (95%, 80%, 70%). Pre-treatment for maximize the exposition of antigen epitopes was achieve in the microwave oven, (2 cycles of 5 minutes each, at 750 Watts), with the slides totally immersed in citrate buffer (pH=6.0). Afterwards, endogenous peroxidases were inhibited with hydrogen peroxide at 3% during 30 minutes, followed by a passage in PBS with detergent, for 5 minutes. The objective of this step was to avoid a possible unspecific reaction of the cromogenic agent, DAB, with other peroxidases, besides those of the complex streptavidinbiotin peroxidase. Then, the cytologies were incubated with Ultra V Block® to reduce the unspecific reaction of the primary antibody with the endogenous avidine and biotine of the cytologies. Subsequently, slides were incubate with the primary HSP70 polyclonal antibody (clone Ab31010; AbCam, UK®), diluted at 1:150, in a humid horizontal chamber, overnight (ca. 18 hours) in a 4ºC temperature. Thereafter, the slides were incubated with Biotinylated Goat AntiPolyvalent® followed by incubation with Streptavidin Peroxidase®. The reaction was visualized through a solution of tetra-hidrocloret 3,3’-diaminobenzidine (DAB; SIGMA®) for 7 minutes and counterstained with Gill’s Hematoxilin . Hematoxilin is a basic nuclear stain that colors the nuclei of the cells in blue, allowing the evidence of the brown color caused by the immunocytochemical markers. The slides were then dehydrated in ethanol of increasing concentrations (95%, 95%, 100% and 100%), diaphanized with xylol and mounted using a synthetic resin, Entellan (Merck®). The results from the immunostaining for HSP70 were evaluated under a magnification lens of 100x. A total of 200 spermatozoa were evaluated for determing both the intensity of the immunoreaction and the sub-celular pattern of immunostaining. The intensity of immunostaining was scored according to a three-point scale (mild = 1, moderate = 2 and strong = 3) or as negative. According to available information in other species, the normal pattern of HSP70 location is the acrosome area. As this molecule is associated with the acrosome envelope, reinforcement of the intensity of immunostaining in the areas not occupied by the nucleus gives to the labeling the appearance of a halo around the spz head. Thus, the immunostaining was evaluated and scored as follows: 40 0 – negative – absence of immunostaining in the acrosome area 1 – The halo is observed but the acrosome membrane presented a faint intensity over the nucleus 2 - The acrossome membrane over the nucleus showed a finely granular immunostaining of moderate intensity, with the halo reinforcement well marked 3 – The acrosome membrane over the nuclear area shows strong reaction, with a dense granular pattern, and also the halo is strongly marked. Further, changes in the subcellular location of the immunostaining were recorded as being dislocated to the neck, middle piece or tail, which included the intermediate and terminal piece of the spermatozoon. In the flagellum and neck, the intensity of immunostaining was not quantified, as it was not possible to distinguish by visualization, differences in the staining intensity. 3.2.4 Statistical analysis Statistical comparisons were performed by using the IBM SPSS Statistics Base 19.0 software for Windows®. Statistical analysis of the effect of treatment over the intensity and sub-celular location of the immunoexpression for HSP70 was performed using the chi-square and Fisher exact tests. A p value ≤ 0.05 was regarded as statistically significant. 3.2.5 Results In all the samples used, HSP70 immunoreaction was found in the majority of sperm cells analysed according to the expected acrosomal pattern. This pattern was observed in more than 98% of the cells in control samples, which only rarely showed dislocation of HSP70 immunolabeling to the tail of the cell. Never dislocation of the immunoreaction into the sperm neck was recorded in control samples. Further, in these samples the intensity score 2 prevailed over all other intensities scores (Figure 15). 41 Figure 15 – Immunocytochemical evidence of HSP70 in the spermatozoa head from the control group spermatozoa. On the left, the presence of positive immunolabeling was visible for the large majority of cells present (Scale bar = 30 m). In the right, in higher magnifications it was clearly visible that the immonoreaction was limited to the acrosome area, whilst the flagellum remained negative for this molecule (Scale bar = 10 m). The normal features for HSP70 protein expression, as determined in this study, were the predominance of moderate intensity of immunoreaction that was exclusively observed over the acrosome membrane. A change in the percentage of exhibition of score 2 and the dislocation of the immunoreaction towards the flagellum was considered as corresponding to a reaction to the treatment. Among our control samples, small individual differences were observed, with samples from ELTON and ELIOS showing slightly higher intensities of immunoreaction than those of the other males (Graphs 7 and 8; Table 9) Overall intensities and pattern of immunolabeling tend to be maintained in treatment A/time 3 groups (Graphs 7 and 8; Table 9), although DINGO´s sample evidence a marked increase in the amount of HSP70 immunoreaction in the sperm cell tail, clearly distinguishing this sample from the cohorts (Figure 16). In contrast to the observed in samples from Treatment A, the samples submitted to freezing/thawed procedures showed an important decreased in the intensity of immunolabeling, along to a dislocation of the immunoreaction towards the sperm tail (Graphs 7 and 8; Table 9) those changes were particularly notorious in DINGO´sample, but also ELTON´s and ELIOS´. The smallest changes were observed for UROCH and ELWOOD´samples (Figure 17). 42 3 2 2 2 1 Figure 16 – Immunocytochemical evidence of HSP70 in chilled spermatozoa (Scale bar = 10m). On the left, smaller magnification. On the right, examples of the three intensity scores for the Spz head used in this study: 1- weak, 2- moderate and 3- strong. Figure 17 - Immunocytochemical staining for HSP70 in frozen/thawed samples (Scale bar = 10 m). On the left, an increased number of cells with weak intensity were found.On the right, besides the different intensities of immunoreaction for the Spz head, we also found a clear dislocation of the immunoreactions for HSP70 into the sperm tail (arrow). The cold treatment significantly influenced the immunoreactions for HSP70 (p=0.006). Controls did not differed from the treatment group A (p=1.0), but both these groups significantly differed from the treatment B group (p=0.09 and p=0.20 respectively). 43 HSP 70 100% Head 80% level 3 60% level 2 40% level 1 20% level 0 Controls DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD 0% Chilling T3 Freezing Graph 7– Distribution of HSP 70 intensity in the spermatozoa’s head HSP 70 100% 80% Tail 60% neg 40% pos 20% Controls Chilling T3 Graph 8 – Distribution of HSP 70 in the spermatozoa’s tail 44 DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD DPART DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD 0% Freezing Table 6 – Immunocytochemical classification of the HSP70 in the spermatozoa Animals DPAR DLIT ELTON ELIOS TECH DINGO UROCH ELWOOD Group Control Treatment A Treatment B Control Treatment A Treatment B Control Treatment A Treatment B Control Treatment A Treatment B Control Treatment A Treatment B control ControlA Treatment B Control Treatment A Treatment B Control Treatment A Treatment B 0 2 2 4 4 6 4 5 2 3 2 3 5 2 3 Degree Head 1 2 21 174 24 170 41 153 16 178 17 172 39 155 4 189 13 181 62 131 4 190 10 182 50 145 14 183 15 180 3 3 4 2 2 5 2 2 4 4 4 5 0 1 2 5 0 6 5 42 8 15 135 145 188 179 52 14 9 5 14 3 3 0 2 25 9 8 17 Neck Tail Tail 2 6 86 7 4 73 5 0 84 7 4 50 3 2 0 2 2 0 0 0 0 0 0 0 0 0 0 0 Midpiece 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Negative 198 192 112 193 196 127 195 200 116 193 196 150 197 198 8 4 0 8 0 0 0 0 0 0 0 0 59 1 27 158 141 199 173 42 178 176 3 1 1 0 0 0 6 6 193 194 170 185 190 180 2 3 2 1 0 0 0 0 0 0 0 0 19 5 3 13 180 195 197 187 4. Discussion Exposition of semen to cold temperatures, in particular to freezing, may induce some changes in the fertility of treated spermatozoa that may compromise the fertility potential of sperm (Quintela, 2010). In this study we compared the behavior of canine sperm samples submitted to chilling and to freezing/thawing procedures with its own control, the freshly ejaculated semen. In the experiment 1 assessment of the sperm cells quality was performed according to classical and advanced technologies, such as CASA and HOST. In experiment 2 we evaluated the changes in HSP70 expression associated to the cold temperature treatments, as HSP may be involved in the cell response to temperature stressors. For all the animals with exception to DINGO, the samples from freshly ejaculated semen used in this work were within the normal range of values accepted for the species. The collected 45 volume was variable within the stud males used, which was probably related to the volume of the 3rd fraction that was collected. Also, the color, pH and morphology of the samples confirmed that they were normal and suitable for this study. Only DINGO revealed some parameters that could compromise its fertility, particularly if a reproductive technology such as artificial insemination was intent to be used. In the case of this male, a large number of abnormal sperm forms were found, along with worsen motility traits and HOST results. Nonetheless, individual variations on the concentration were observed. This in fact agrees with previous studies that demonstrated that individual variations exist in the sperm concentration between males and also within collections from the same male (Johnston et al., 2001). CASA analysis is now currently used in larger artificial insemination centers, as it allows a less subjective evaluation of sperm motility and velocity. Assessment of sperm motility is not a direct measure of the ratios between the number of dead and live spermatozoa, but it gives some information about its fertilizing ability as it reflects its structural and functional competence (Peña Martinez, 2004). Though the predictive value of sperm morphology in determining pregnancy rates is low, it is recognized that sperm motility, particularly sperm progressive motility and speed are important parameters for sperm fertility. The most regarded parameter to evaluate the semen quality is the motility, however, very frequently the motility parameters are directly linked to the increase of abnormal Spz forms (Rijsselaere et al., 2004). In control samples it was also found the existence of small differences concerning the total and progressive motility, however not compromising the quality of the samples, taking into account that most of our samples presented what is considered a good quality semen, over 70% in total motility (Feldman et al., 2004). Only DINGO´s control sample showed slightly lower values for motility in comparison to those of the other males. When chilled, in treatment A, at the times 1, 2 and 3, spermatozoa showed a decrease in the total and progressive motility for 88% of our sample population. This is in concordance with other published works, and is considered to be normal and usual. The values found in the study presented here are within the average for the species (Rota et al., 1995; Bessa, 2005). In contrast, in samples submitted to treatment B, the results for both progressive and total motility show an important decrease. Although it coincides with the average described in other published data, one animal in our experiments showed a very bad performance under freezing conditions (DINGO), 46 evidencing a huge decrease in motility. Such an event might allow the classification of this male as a “bad freezer” (England et al., 1996; Bessa, 2005), even if it was the sole defect found in this male. However, DINGO also presented other characteristics, such as the increased number of abnormal sperm cells, which could be related to the low motility parameters and lower number of rapid cells whilst the static Spz increased particular after treatment. The hypo-osmotic swelling test (HOST) allows the identification of spermatozoa which possess a functional intact membrane. The integrity of the sperm cell membrane is of crucial importance, as it allows maintaining both biochemical and structural integrity of the spermatozoa and is essential to gamete interaction at fertilization (Quintela, 2010). When exposed to hypoosmotic solutions, biochemically-active, and membrane intact spermatozoa increase their volume in order to re-establish the equilibrium between the fluid compartment within the spermatozoa and the extracellular environment. Hypo-osmotic induced cellular swelling implies changes in both cell size and shape that can be visualized in a phase contrast microscope. The swelling process culminates with the spherical expansion of the cell membrane covering the tail that forces the flagellum to coil inside the membrane (Quintela, 2010). When evaluating the cold treatment effects on sample responses to the hypo-osmotic swelling test, it was found a slight decrease on the number of coiled spermatozoa’s tails for treatment A, although all the samples maintained acceptable values, comparable to those reported in other available studies (Rota et al., 1995). In contrast, for treatment B, a significant decrease in the coiled spermatozoa was observed for all the samples, which was more pronounced for DINGO´s sample. This corresponds to a loss of membrane integrity in frozen/thawed spermatozoa, which has been described for the dog in the available literature (England et al., 1996; Pinto et al., 2008). According to the evaluation of morphology, the CASA parameters and the HOST results, we could categorize the animals according to levels of quality. Crossing information from all those tests, in particular comparing reduction in parameters such as progressive motility and increase in membrane damaged spermatozoa, we propose the classification of the animals as follow: Excellent freezers, where DPAR, ELTON and ELWOOD would be included; Good freezers, which would include ELIOS and UROCH; Acceptable freezers were DLIT and TECH would be included; 47 We could consider DINGO as being a bad freezer, but indeed, DINGO´samples presented some bad characteristics since the beginning. To be sure, we should perform additional tests in this animal to ascertain the existence of an eventual sub-clinical concurrent disease that could interfere with the male fertility, or a complete breeding examination to determine its (in)fertility level. The heat-shock response is a highly conserved mechanism in all the organisms, which may be mediated by extreme proteotoxic insults, such as heat, oxidative stress, heavy metals, toxins or even bacterial infections (Åkerfelt 2010). It is now believed that heat-shock response is essential for survival in a stressful environment. Spermatozoa’ ability to respond to osmotic, oxidative and cryopreservation induced cell stress is not completely well understood. The sperm subpopulations survival at cryopreservation, within a single ejaculate may reflect different maturational states, the activation of cell protective mechanisms or status of membrane stability at the time of cryopreservation. In order to improve cryopreservation protocols, it is critical to define the characteristics of surviving sperm in order to minimize the negative effects of cryopreservation on functions important to sperm function. In controls, a moderate HSP70 immunoreaction was found in the sperm head, at the acrosomal level. In freshly ejaculated spermatozoa, tail immunostaining was seldom observed. This was also found in other species, such as the pig and bull, but also for the dog (Volpe et al, 2008). In cold temperature-stressed samples, some changes in the intensity of immunoreactions and more or less notorious dislocation of the immunostaining into the flagellum. These changes were less pronounced in chilled samples (treatment A) than in freeze/thawing samples (treatment B). This sort of modification in the sub-celular location of the immunostaining that co-existed, in our study, with a decrease in the intensity score for acrosomal staining has been described in the pig as being a “capacitated” pattern of HSP70 immunoexpression. Considering that the variation in the pattern for HSP immunostaining is not equal to all the animals, we should suggest that it could be associated also with different ability to resist to cold-induced lesions. In fact, the case with poor fertility potential (DINGO) was also the one showing the higher decreases in the intensity scores in the acrosome and staining dislocation after treatment B. Those changes occurred in parallel with reduction in motility and a decrease in the HOST test. It is possible that those events may be associated. In fact, as spermatozoa has a 48 limited synthetic ability, under stressing conditions the cell is much dependent on the ability of activating the defense mechanisms available. During the cryopreservation of sperm, both osmotic and oxidative stressors are thought to contribute to membrane and cellular damage which leads to the reduction in post-thaw motility and viability (McCarthy, et al., 2010). As known effects of osmotic stress on the sperm cell, there is the production of oxygen free radicals, leading to the peroxidation of the membrane lipids (Alvarez et al., 1992; McCarthy et al., 2010), a decrease in progressive and total motility, an increase in the tyrosine phosphorylation of tail proteins (Liu et al., 2006) and a change in the actin cytoskeleton (Correa et al., 2007). Spermatozoa are sensible to various kinds of stresses, such as, the oxidative stress, once they lack cytoplasmatic defenses (Saleh et al., 2002). The sperm plasma membrane is known to contain lipids in the form of polyunsaturated fatty acids, which are vulnerable to ROS (reactive oxygen species), that together with the lipids, triggers a chain of chemical reactions which is called lipid peroxidation (Eddy, 1999). Being aware that the heat-shock response is a highly conserved mechanism in all the organisms, which is induced by extreme proteotoxic insults, such as heat, oxidative stress, heavy metals, toxins or even bacterial infections, it is believed that this heat-shock response is essential for survival in a stressful environment. Spermatozoa’s ability to respond to osmotic, oxidative and cryopreservation induced cell stress is not completely well understood. The sperm subpopulations survival at cryopreservation, within a single ejaculate may reflect different maturational states, the activation of cell protective mechanisms or status of membrane stability at the time of cryopreservation. In order to improve cryopreservation protocols, it is critical to define the characteristics of surviving sperm in order to minimize the negative effects of cryopreservation on functions important to sperm maturation. In our study, after the cryopreservation procedure, more exclusively the chilling part, we could observe a slight decrease on the marking intensity of the heat shock proteins 70 in the head of the spermatozoa’s in a general point of view, which correlating with the increase noted in the presence of the HSP in the tail, can evidence the dislocation of them in response to a stress factor caused by the cryopreservation procedure. This dislocation is far more evident, when we evaluate the immunocytochemistry of the frozen samples. All the animals show a relevant decrease of the level 2 head marking, an increase on the level 1 marking, noting a reduction of the activity or amount of HSP70 on the acrosome/head of the spermatozoa and consequently an increase in the amount/activity of the HSP70 on the tail region. Considering that the variation is 49 not equal to all the animals we should take into account a great number of variables and limitations to evaluate the global equation. During the cryopreservation of sperm, both osmotic and oxidative stressors are thought to contribute to membrane and cellular damage which leads to the reduction in post-thaw motility and viability (McCarthy, et al., 2010). 50 5. Final considerations As find by other authors, we observed that the preservation processes may cause a decrease in the quality of the dog semen. This loss of quality parameters is far more evidenced in the freezing procedure, rather than in the chilling one, making it important to further study the reasons, in order to improve the methods of cryopreservation. The heat-shock induces significant changes in the viability, motility and consequently in the fertilizing capacity of the dog spermatozoa. Heat stress causes an increase in the expression of the HSP70 and a dislocation of these to the caudal part of the spermatozoa, specifically to the tail of the spermatozoa. There is a diferential increase in the expression of the HSP70 under different cold treatments, resulting that in the freezing procedure, the described above is strongly observed, rather than in the chilling procedure, where subtle changes are duly noted. Further experiments are being undertaken to evaluate better the distribution and role of the HSP70, along with other HSP, in order to develop cryopreservation protocols which promote cell survival. 51 References Åkerfelt RIMaLS. Heat shock factors: integrators of cell stress, development and lifespan. Nature Reviews Molecular Cell Biology. 2010; (11): 545–555. Althouse GC, Ko JC, et al. Effect of latex and vinyl examination gloves on canine spermatozoal motility. J Am Vet Med Assoc. 1991; 199 (2): 227-229. Alvarez JG and Storey BT. Evidence for increased lipid peroxidative damage and loss of superoxide dismutase activity as a mode of sublethal cryodamage to human sperm during cryopreservation. J Androl. 1992; 13 (3): 232-241. Andersen K. Fertility of Frozen Dog Semen. Acta Veterinaria Scandinavica. 1972; 13 (1): 128134. Baker HW and Clarke GN. Sperm morphology: consistency of assessment of the same sperm by different observers. Clin Reprod Fertil. 1987; 5 (1-2): 37-43. Bangham AD and Hancock JL. A new method for counting live and dead bull spermatozoa. Nature. 1955; 176 (4483): 656. Bencharif D, Amirat L, et al. The advantages of LDL (low density lipoproteins) in the cryopreservation of canine semen. Theriogenology. 2008; 70 (9): 1478-1488. Bessa ACAM.. Influencia en la calidad espermática de la adición de distintas concentraciones de crioprotectores para la conservación del semen canino. Departamento de Medicina y Cirugía Animal. (2005); Tesis (Doctoral). Boucher JH, Foote RH, et al. The evaluation of semen quality in the dog and the effects of frequency of ejaculation upon semen quality, libido, and depletion of sperm reserves. Cornell Vet. 1958; 48 (1): 67-86. Burgess CM, Bredl JCS, et al. Vital and ultrastructural changes in dog spermatozoa during cryopreservation. Advances in Reproduction in Dogs, Cats and Exotic Carnivores. 2001; (57): 357-363. CBRA CBdRA. Manual para exame andrológico e avaliação de sêmen animal, 2nd ed. 1998. Chong AP, Walters CA, et al. The neglected laboratory test. The semen analysis. J Androl. 1983; 4 (4): 280-282. Correa LM, Thomas A, et al. The macaque sperm actin cytoskeleton reorganizes in response to osmotic stress and contributes to morphological defects and decreased motility. Biol Reprod. 2007; 77 (6): 942-953. Craig EA. The heat shock response. CRC Crit Rev Biochem. 1985; 18 (3): 239-280. Cunningham JG and Klein BG, Textbook of veterinary physiology. In: Book Textbook of veterinary physiology, Editor., Saunders, an imprint of Elsevier Inc. 2007. Dott HM and Foster GC. A technique for studying the morphology of mammalian spermatozoa which are eosinophilic in a differential 'life-dead' stain. J Reprod Fertil. 1972; 29 (3): 443-445. Drobnis EZ, Crowe LM, et al. Cold shock damage is due to lipid phase transitions in cell membranes: a demonstration using sperm as a model. J Exp Zool. 1993; 265 (4): 432437. Eddy EM. Role of heat shock protein HSP70-2 in spermatogenesis. Rev Reprod. 1999; 4 (1): 2330. 52 England G and Heimendahl A, BSAVA Manual of Canine and Feline Reproduction and Neonatology In: Book BSAVA Manual of Canine and Feline Reproduction and Neonatology Editor. 2nd edition. 2010: 18-22;70-78. England GC. Semen quality in dogs and the influence of a short-interval second ejaculation. Theriogenology. 1999; 52 (6): 981-986. England GC and Allen WE. Evaluation of cellulose acetate/nitrate filters for measuring the motility of dog spermatozoa. J Reprod Fertil. 1990; 88 (1): 369-374. England GC and Plummer JM. Hypo-osmotic swelling of dog spermatozoa. J Reprod Fertil Suppl. 1993; 47: 261-270. England GCW and Allen WE. Factors Affecting the Viability of Canine Spermatozoa .1. Potential Influences during Processing for Artificial-Insemination. Theriogenology. 1992; 37 (2): 363-371. England GCW, Burgess CM, et al. Relationship between the fertile period and sperm transport in the bitch. Theriogenology. 2006; 66 (6-7): 1410-1418. England GCW and Ponzio P. Comparison of the quality of frozen-thawed and cooled-rewarmed dog semen. Theriogenology. 1996; 46 (1): 165-171. Ettinger SJ and Feldman EC, Textbook of veterinary internal medicine : diseases of the dog and the cat. In: Book Textbook of veterinary internal medicine : diseases of the dog and the cat, Editor. 7th ed. St. Louis, Mo., Elsevier Saunders. 2010. Farstad W. Cryopreservation of canine semen - new challenges. Reproduction in Domestic Animals. 2009; 44 Suppl 2: 336-341. Feldman EC and Nelson RW. Clinical and diagnostic evaluation of the male reproductive tract. In: Canine and Feline Endocrinology and Reproduction. Philadelphia, W. B. Saunders. 1996: 673-690. Feldman EC and Nelson RW, Canine and feline endocrinology and reproduction. In: Book Canine and feline endocrinology and reproduction, Editor. 3rd ed. St. Louis, Mo., Saunders. 2004. Foote RH. The history of artificial insemination: Selected notes and notables. Journal of Animal Science. 2002; 80 (E-Suppl 2): 1-10. Freshman JL. Clinical management of the subfertile stud dog. Vet Clin North Am Small Anim Pract. 2001; 31 (2): 259-269. Freshman JL. Semen collection and evaluation. Clin Tech Small Anim Pract. 2002; 17 (3): 104107. Froman DP and Amann RP. Inhibition of motility of bovine, canine and equine spermatozoa by artificial vagina lubricants. Theriogenology. 1983; 20 (3): 357-361. Georgopoulos C and Welch WJ. Role of the major heat shock proteins as molecular chaperones. Annu Rev Cell Biol. 1993; 9: 601-634. Gordon I. Reproductive technologies in farm animals. Wallingford, Oxfordshire, UK, Cambridge, MA, CABI Pub. 2004. Gunn RMC. Fertility in sheep; artificial production of seminal ejaculation and the characters of the spermatozoa contained there, in. Melbourne,, H.J. Green. 1936. Gunzel-Apel AR, Gunther C, et al. Computer-assisted analysis of motility, velocity and linearity of dog spermatozoa. J Reprod Fertil Suppl. 1993; 47: 271-278. 53 Hay MA, King WA, et al. Effects of cooling, freezing and glycerol on penetration of oocytes by spermatozoa in dogs. J Reprod Fertil Suppl. 1997; 51: 99-108. Hermansson U. Studies of canine and feline sperm viability under different storage procedures. Diss. (sammanfattning/summary) Uppsala : Sveriges lantbruksuniv., Acta Universitatis agriculturae Sueciae, 1652-6880 ; 2006:68. 2006. Hewitt DA and England GC. The canine oocyte penetration assay; its use as an indicator of dog spermatozoal performance in vitro. Anim Reprod Sci. 1998; 50 (1-2): 123-139. Hewitt DA and England GCW. Manipulation of canine fertility using in vitro culture techniques. Advances in Reproduction in Dogs, Cats and Exotic Carnivores. 2001; (57): 111-125. Iguer-Ouada M and Verstegen JP. Long-term preservation of chilled canine semen: effect of commercial and laboratory prepared extenders. Theriogenology. 2001; 55 (2): 671-684. Iguer-Ouada M and Verstegen JP. Evaluation of the "Hamilton Thorn computer-based automated system" for dog semen analysis. Theriogenology. 2001a; 55 (3): 733-749. Iguer-Ouada M and Verstegen JP. Validation of the sperm quality analyzer (SQA) for dog sperm analysis. Theriogenology. 2001b; 55 (5): 1143-1158. Jequier AM and Ukombe EB. Errors inherent in the performance of a routine semen analysis. Br J Urol. 1983; 55 (4): 434-436. Jeyendran RS, Van der Ven HH, et al. Development of an assay to assess the functional integrity of the human sperm membrane and its relationship to other semen characteristics. J Reprod Fertil. 1984; 70 (1): 219-228. Johnson C. Current concepts on infertility in the dog. Waltham Focus Vol 16 No 2. 2006. Johnston SD. New canine semen evaluation techniques for the small animal practitioner. In 1989 Proceedings of the Annual Meeting, Society for Theriogenology, Coeur d'Alene, ID." 320-326. 1989. Johnston SD. Performing a complete canine semen evaluation in a small animal hospital. Vet Clin North Am Small Anim Pract. 1991; 21 (3): 545-551. Johnston SD, Root Kustritz MV, et al., Canine and feline theriogenology. In: Book Canine and feline theriogenology, Editor. 1st ed. Philadelphia, PA, Saunders. 2001. Juniewicz PE, Lemp BM, et al. Transrectal ultrasonography as a method to monitor canine prostatic size in situ: measurements following endocrine manipulation and ejaculation. Prostate. 1989; 14 (3): 265-277. Kruger TF, Dutoit TC, et al. Sperm Morphology - Assessing the Agreement between the Manual Method (Strict Criteria) and the Sperm Morphology Analyzer Ivos. Fertility and Sterility. 1995; 63 (1): 134-141. Kumi-Diaka J. Subjecting canine semen to the hypo-osmotic test. Theriogenology 39, 1279– 1289. 1993. Kustritz MVR. The value of canine semen evaluation for practitioners. Theriogenology. 2007; 68 (3): 329-337. Kutzler MA. Semen collection in the dog. Theriogenology. 2005; 64 (3): 747-754. Kutzler MA, Solter PF, et al. Characterization and localization of alkaline phosphatase in canine seminal plasma and gonadal tissues. Theriogenology. 2003; 60 (2): 299-306. Lagishetty V, Liu NQ, et al. Vitamin D metabolism and innate immunity. Mol Cell Endocrinol. 2011. 54 Larsson B and Rodriguez-Martinez H. Can we use in vitro fertilization tests to predict semen fertility? Anim Reprod Sci. 2000; 60-61: 327-336. Lawson R, Mestril R, et al. Expression of heat shock-beta-galactosidase hybrid genes in cultured Drosophila cells. Mol Gen Genet. 1984; 198 (1): 116-124. LeFrapper L, Walston, L., & Whisnant, C. S. Comparison of various extenders for storage of cooled stallion spermatozoa for 72 hours. Journal of Equine Veterinary Science. 2010; 30 (4): 200-204. Linde-Forsberg C, Strom Holst B, et al. Comparison of fertility data from vaginal vs intrauterine insemination of frozen-thawed dog semen: a retrospective study. Theriogenology. 1999; 52 (1): 11-23. Liu DY, Clarke GN, et al. Hyper-osmotic condition enhances protein tyrosine phosphorylation and zona pellucida binding capacity of human sperm. Hum Reprod. 2006; 21 (3): 745752. Luvoni GC. Gamete cryopreservation in the domestic cat. Theriogenology. 2006; 66 (1): 101111. Martinez AI. Canine fresh and cryopreserved semen evaluation. Anim Reprod Sci. 2004; 82-83: 209-224. McCarthy MJ, Baumber J, et al. Osmotic stress induces oxidative cell damage to rhesus macaque spermatozoa. Biol Reprod. 2010; 82 (3): 644-651. Monteiro JC, Goncalves JS, et al. Influence of ascorbic acid and glutathione antioxidants on frozen-thawed canine semen. Reproduction in Domestic Animals. 2009; 44 Suppl 2: 359362. Mortimer D, Shu MA, et al. Standardization and quality control of sperm concentration and sperm motility counts in semen analysis. Hum Reprod. 1986; 1 (5): 299-303. Nelson RW and Couto CG, Small animal internal medicine. In: Book Small animal internal medicine, Editor. 4th ed. St. Louis, Mo., Mosby/Elsevier. 2009. Nollen EA, Brunsting JF, et al. In vivo chaperone activity of heat shock protein 70 and thermotolerance. Mol Cell Biol. 1999; 19 (3): 2069-2079. Nöthling JO and Shuttleworth R. The effect of straw size, freezing rate and thawing rate upon post-thaw quality of dog semen. Theriogenology. 2005; 63 (5): 1469-1480. Oettle EE. Changes in Acrosome Morphology during Cooling and Freezing of Dog Semen. Animal Reproduction Science. 1986; 12 (2): 145-150. Oettle EE. Sperm morphology and fertility in the dog. J Reprod Fertil Suppl. 1993; 47: 257-260. Oettlé EE. Changes in acrosome morphology during cooling and freezing of dog semen. Animal Reproduction Science. 1986; 12 (2): 145-150. Parcellier A, Gurbuxani S, et al. Heat shock proteins, cellular chaperones that modulate mitochondrial cell death pathways. Biochem Biophys Res Commun. 2003; 304 (3): 505512. Pena A, Johannisson A, et al. Post-thaw evaluation of dog spermatozoa using new triple fluorescent staining and flow cytometry. Theriogenology. 1999; 52 (6): 965-980. Peña A and Linde-Forsberg C. Effects of Equex, one- or two-step dilution, and two freezing and thawing rates on post-thaw survival of dog spermatozoa. Theriogenology. 2000; 54 (6): 859-875. 55 Pena AI, Quintela LA, et al. Viability assessment of dog spermatozoa using flow cytometry. Theriogenology. 1998; 50 (8): 1211-1220. Pena AI, Quintela LA, et al. Flow cytometric assessment of acrosomal status and viability of dog spermatozoa. Reproduction in Domestic Animals. 1999; 34 (6): 495-502. Pinto CR and Kozink DM. Simplified hypoosmotic swelling testing (HOST) of fresh and frozenthawed canine spermatozoa. Anim Reprod Sci. 2008; 104 (2-4): 450-455. Pinto CRF and Kozink DM. Simplified hypoosmotic swelling testing (HOST) of fresh and frozen-thawed canine spermatozoa. Animal Reproduction Science. 2008; 104 (2-4): 450455. Purswell BJA, G. and Root MV. Guidelines for Using the Canine Breeding Soundness Evaluation Form. Theriogenology Handbook. Society for Theriogenology. 1992. Quinn PJ and White IG. The effect of cold shock and deep-freezing on the concentration of major cations in spermatozoa. J Reprod Fertil. 1966; 12 (2): 263-270. Quintela IRSO, A.O. Souza, A.L. Gusmão, A.R. Silva. Water-induced hypo-osmotic test for the evaluation of canine sperm membrane integrity. Animal Reproduction Science. 2010; 7 (2). Rijsselaere T, Van Soom A, et al. Automated sperm morphometry and morphology analysis of canine semen by the Hamilton-Thorne analyser. Theriogenology. 2004; 62 (7): 12921306. Rijsselaere T, Van Soom A, et al. New techniques for the assessment of canine semen quality: a review. Theriogenology. 2005; 64 (3): 706-719. Risopatron J, Catalan S, et al. Effect of albumin and polyvinyl alcohol on the vitality, motility and acrosomal integrity of canine spermatozoa incubated in vitro. Reproduction in Domestic Animals. 2002; 37 (6): 347-351. Rodríguez-Gil JE, Montserrat A, et al. Effects of hypoosmotic incubation on acrosome and tail structure on canine spermatozoa. Theriogenology. 1994; 42 (5): 815-829. Rodríguez-Martínez H. Evaluation of frozen semen: traditional and new approaches. Chenoweth PJ, Topics in bull fertility. Ithaca NY: International Veterinary Information Service. (http://www.ivis.org). 2000: 502-600. Rota A, Pena AI, et al. In vitro capacitation of fresh, chilled and frozen-thawed dog spermatozoa assessed by the chloretetracycline assay and changes in motility patterns. Anim Reprod Sci. 1999; 57 (3-4): 199-215. Rota A, Strom B, et al. Effects of seminal plasma and three extenders on canine semen stored at 4 degrees C. Theriogenology. 1995; 44 (6): 885-900. Saleh RA and Agarwal A. Oxidative stress and male infertility: from research bench to clinical practice. J Androl. 2002; 23 (6): 737-752. Schafer-Somi S and Aurich C. Use of a new computer-assisted sperm analyzer for the assessment of motility and viability of dog spermatozoa and evaluation of four different semen extenders for predilution. Anim Reprod Sci. 2007; 102 (1-2): 1-13. Schlesinger MJ. Heat shock proteins: the search for functions. J Cell Biol. 1986; 103 (2): 321325. 56 Schlesinger MJ, Ashburner M, et al., Heat shock, from bacteria to man. In: Book Heat shock, from bacteria to man, Editor. ed. Cold Spring Harbor, N.Y., Cold Spring Harbor Laboratory. 1982. Schubert CL and Seager SWJ. Semen Collection and Evaluation for the Assessment of Fertility Parameters in the Male Dalmatian. Canine Practice. 1991; 16 (5): 17-21. Seager SWJ. Successful pregnancies utilizing frozen dog semen. A.I. Digest 17,6,16. 1969. Seager SWJ. Artificial insemination in dogs. Em: Small Animal Reproduction and Infertility: A Clinical Approach to Diagnosis and Treatment, Ed. T.J. Burke. Lea & Febiger, Philadelphia. 1986: 207-210. Silva AR, Cardoso RCS, et al. Quality of canine semen submitted to single or fractionated glycerol addition during the freezing process. Theriogenology. 2003; 59 (3-4): 821-829. Simpson GM, England GCW, et al., BSAVA manual of small animal reproduction and neonatology. In: Book BSAVA manual of small animal reproduction and neonatology, Editor., British Small Animal Veterinary Association. 1998. Smith SC and England GC. Effect of technical settings and semen handling upon motility characteristics of dog spermatozoa measured using computer-aided sperm analysis. J Reprod Fertil Suppl. 2001; 57: 151-159. Sonna LA, Fujita J, et al. Invited review: Effects of heat and cold stress on mammalian gene expression. J Appl Physiol. 2002; 92 (4): 1725-1742. Strom B, Rota A, et al. In vitro characteristics of canine spermatozoa subjected to two methods of cryopreservation. Theriogenology. 1997; 48 (2): 247-256. Strom Holst B, Larsson B, et al. Evaluation of chilled and frozen-thawed canine spermatozoa using a zona pellucida binding assay. J Reprod Fertil. 2000a; 119 (2): 201-206. Strom Holst B, Larsson B, et al. Sperm binding capacity and ultrastructure of the zona pellucida of stored canine oocytes. J Reprod Fertil. 2000b; 119 (1): 77-83. Ström Holst RA, Andersen Berg K, Linde-Forsberg C and Rodriguez-Martinez H. Canine sperm damage after freezing–thawing: ultrastructural evaluation and content of selected elements. Reproduction in Domestic Animals. 1998; 33: 77-82. Swanson WF. Application of assisted reproduction for population management in felids: the potential and reality for conservation of small cats. Theriogenology. 2006; 66 (1): 49-58. Thomassen R and Farstad W. Artificial insemination in canids: A useful tool in breeding and conservation. Theriogenology. 2009; 71 (1): 190-199. Verstegen J, Iguer-Ouada M, et al. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology. 2002; 57 (1): 149-179. Volpe S, Leoci R, et al. Relationship between motility and mitochondrial functional status in canine spermatozoa. Reproduction in Domestic Animals. 2009; 44 Suppl 2: 275-278. Wanke MM. Canine brucellosis. Animal Reproduction Science. 2004; 82-83: 195-207. Watson PF. The effects of cold shock on sperm cell membranes. Academic Press, New York. 1981: 189-218. Watson PF. The causes of reduced fertility with cryopreserved semen. Anim Reprod Sci. 2000; 60-61: 481-492. White IG. Lipids and calcium uptake of sperm in relation to cold shock and preservation: a review. Reprod Fertil Dev. 1993; 5 (6): 639-658. 57 WHO, WHO laboratory manual for the examination and processing of human semen. In: Book WHO laboratory manual for the examination and processing of human semen, Editor. 5th ed. Geneva, World Health Organization. 2010. Wilmot S. From public service' to artificial insemination: animal breeding science and reproductive research in early twentieth-century Britain. Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences. 2007; 38 (2): 411-441. Woelders H. Fundamentals and recent development in cryopreservation of bull and boar semen. Vet Q. 1997; 19 (3): 135-138. Yildiz C, Kaya A, et al. Influence of sugar supplementation of the extender on motility, viability and acrosomal integrity of dog spermatozoa during freezing. Theriogenology. 2000; 54 (4): 579-585. 58 ANNEXES 59 Annex 1 Total and progressive motility and consequent treatment losses Total Motility (%) losses associated with treatment Treatment A Time 0 (Fresh) Time 1 Time 2 Time 3 DPAR 89 -3 -4 -5 -18 DLIT 85 -19 -18 -16 -32 ELTON 68 18 4 13 -15 ELIOS 80 7 0 8 -25 TECH 89 -13 -8 -4 -39 DINGO 61 21 10 20 -45 UROCH 88 -7 -1 -1 -28 ELWOOD 73 8 4 16 -12 Treatment B Progressive Motility (%) losses associated with treatment Treatment A Time 0 (Fresh) Time 1 Time 2 Time 3 DPAR 61 -12 -9 -2 -22 DLIT 61 -22 -21 -17 -27 ELTON 49 0 4 4 -6 ELIOS 54 -5 -2 -4 -21 TECH 55 -2 -3 -8 -24 DINGO 32 26 18 21 -20 UROCH 52 0 2 -5 -20 ELWOOD 50 1 3 4 -7 60 Treatment B Annex 2 CASA parameters for sperm cells under cold treatments, when compared to the controls (fresh semen). Treatment A – Chilling (times 1, 2 and 3 respectively, 1.5, 4 and 24h of refrigeration); treatment B – Freezing/Thawing. VAP (μm/s) Controls Treatment A Time 1 Time 2 Time 3 Treatment B DPAR 155.5 152.5 153.8 148.7 132.8 DLIT ELTON ELIOS 160.2 137.4 137.3 134.6 142.3 148.5 134 125.7 144.7 133.9 134.4 148.3 115.7 126.2 123 TECH 111.9 148.3 148.8 146.5 125.8 DINGO UROCH ELWOOD 105.7 142.7 139.5 147.8 133.1 150.5 143.9 136 147.7 137.2 140.3 153 111.1 108.3 111.3 DPAR DLIT 130.7 137.6 123.2 111.5 125.9 110.1 124.6 110.8 106.7 99.8 ELTON ELIOS TECH 121.9 116.7 89.7 113.5 117.9 127.9 110.6 120.5 123.7 110 117.4 114 115.2 101.5 107.4 DINGO 83.1 125.7 124.1 112 100.7 UROCH ELWOOD 114.9 119.9 110.1 124.8 110.2 124.9 108.5 122.3 88 97 DPAR 205.8 208.2 213 231.7 183.5 DLIT ELTON 212 167.1 187.2 201.5 209.5 156.7 237.6 175.4 162.3 152.5 ELIOS 177.9 210.7 189 205 172.9 TECH DINGO 186.6 187.7 183.9 195.1 192.6 190.2 209 204.6 161.2 143.9 VSL (μm/s) VCL (μm/s) UROCH 193.9 179.2 185.3 206.1 152.4 ELWOOD 191.2 199 194.4 212.5 159 DPAR 6.8 7.6 7.7 9.1 6.8 ALH (μm) DLIT 7.4 7.6 8.9 10.5 16.6 ELTON 5.9 7.8 6.3 7 5.4 ELIOS 6.5 7.7 6.8 7.3 7.0 TECH 7.7 6.6 7.4 7.9 5.7 DINGO UROCH 8.1 6.5 7.5 4.9 7 7.1 8.1 7.9 6.1 6 ELWOOD 7.3 7.3 7.8 3.3 6.6 61 STR (%) DPAR 83 78 79 83 77 DLIT 85 80 80 81 85 ELTON ELIOS 87 84 78 77 86 81 81 74 90 80 TECH 80 84 81 76 83 DINGO 78 83 84 81 88 UROCH 79 81 79 76 79 ELWOOD 84 80 82 78 86 DPAR DLIT 65 66 60 60 60 54 56 50 59 63 ELTON 73 59 72 65 76 ELIOS 68 58 65 59 60 TECH 50 70 65 56 67 DINGO UROCH 46 62 65 63 66 61 57 55 71 59 ELWOOD 64 63 65 59 62 LIN (%) 62
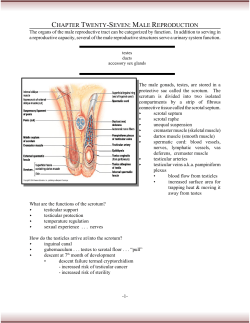









© Copyright 2026