ABC
docz
Explore
Log in
Create new account
Download
Report
health and fitness
disease
cancer
Document 373520
Health Canada Approves ABRAXANE Plus Gemcitabine for First-Line Treatment
Document 148333
MINIREVIEWS
2120 Gastrointestinal Function Chemistries Profile Percentile Ranking by Quintile
INTRODUCTION
Evaluation of Trace Elements in Pancreatic Cancer Patients in Iran Original Article
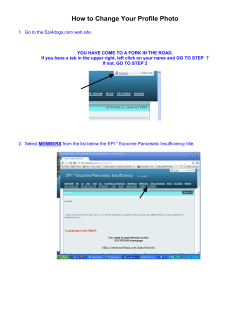
How to Change Your Profile Photo
How to Deal with a Periampullary Mass?
Pancreatic Cancer Therapy: Current Status Timothy R. Donahue, MD
2nd Annual TIMCC Symposium Wednesday October 22 , 2014
© Copyright 2026
About abcdocz
DMCA / GDPR
Report