ABC
docz
Explore
Log in
Create new account
Download
Report
No category
Oseltamivir-Resistant Influenza A (H1N1) Virus Strain with an
Avian Influenza – What is it? Avian influenza
USDA Stakeholders Announcement
Health Education Presentation on Seasonal Influenza 2015
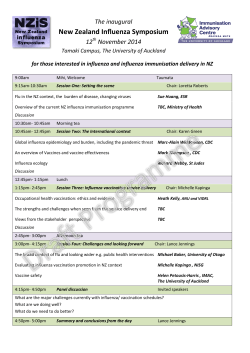
New Zealand Influenza Symposium The inaugural 12 November 2014
Health Education Presentation on Seasonal Influenza 2015
10X Essentials Newsletter - Geisinger Medical Laboratories
Medical Briefings Avian Influenza
Safe Banking Caveats
DOPH Influenza Surveillance Weekly Update 2015 Week 5
BIOSECURITY ALERT
© Copyright 2026
About abcdocz
DMCA / GDPR
Report