
Inheritance of Seed Hardness and Seed Coat Weight in Faba Bean
Inheritance of Seed Hardness and Seed Coat Weight in Faba Bean (Vicia faba L.) A Thesis Submitted to the University of Khartoum in Fulfillment of the Requirements for the Degree of Doctor of Philosophy (Agric.) By Ikram Elfadul Abdalla Eltayb B.Sc.(Agric.) Honours University of Khartoum (May 1987) M.Sc.(Agric.) University of Khartoum (1993) Supervisor : Professor Farouk Ahmed Salih Faculty of Agriculture University of Khartoum Department of Agronomy 2004 DEDICATION To my family : My Husband : Omer Elzein To his continued interest .. My Kids: with a lot love unlimited encouragements SUMMARY AND CONCLUSION 1- The main objective of this research has been to study the magnitude of gene action controlling the inheritance of seed hardness and seed coat weight of faba bean (Vicia faba L.). in F1, F2 and their respective parents. Eight genotypes of faba bean were used as parents in this study. The eight parents were selected in season 2000/2001 according to their variability in seed hardness and seed coat weight. They were divided into tow groups, low hard seeded parents (< 5.0% hard seeds) and high hard seeded parents ( > 12.0% hard seeds). 2- In the first season (2001/2002), the eight parents were crossed in a half diallel way producing 28 F1 hybrids. In the second season (2002/2003), the eight parents, together with the 28 F1 hybrids were grown in a therano cage for selfing and the F2 seeds were obtained. The parents, F1 hybrids and F2 i.e. a total of 64 genotypes, were grown in the third season. A randomized complete block design with two replications was used to test the material. At harvest, in the later two seasons (the evaluation seasons), data were collected, on mean bases, for the following four parameters: 1. number of pods/plant, 2. number of seeds/plant, 3. 100-seed weight(g), and 4. seed yield/plot (g). The percentages of the two seed quality characters, seed hardness and seed coat weight, were determined as described by Salih (1982). 3- The genetical analysis was performed using Hayman (1954, 1958) method to determine the magnitude of gene action controlling the inheritance of seed hardness and seed coat weight. Also Griffing (1956) Method 2, Model 1, was used for combining ability analysis. Both analysis of components of genetic variations and the (wr,vr) graphs revealed seed hardness as a dominant character and the nonadditive gene effects (b, H1 and H2) were more important in the inheritance of this character. Ambidirectional dominance of the character and un equal distribution of genes also were detected . The mean degree of dominance [(H1/D) ½ ratio, and the interception points, a ] indicated an over dominance in the inheritance of seed hardness. Also a predominance of the dominant alleles in the parents used was observed, where positive F and Fri estimates and negative r values were obtained. A group of two to three genes were detected dominant in the inheritance of seed hardness, among them a single gene was detected. Regarding the inheritance of seed coat weight, the results revealed that the character was under the control of both additive and nonadditive genotypic gene action. ratio (H1) However, the dominant component of the was more important. Ambidirectional dominance of the character was observed (b1 is not significant) and the non-additive genetic components , H2, b2, were significant in both F1 and F2 indicating unequal distribution of genes in the parents used. Positive F estimates were obtained in the two generations indicating that, dominant alleles were more frequent than recessive ones. Three to four genes were found to control seed coat weight indicating its quantitative inheritance. Among them one gene was dominant. Partial dominance was shown by F2 and F1 in the 2nd evaluation season (F1 2004) , while F1 in the first evaluation season (F1 2003) exhibited over dominance. In the (wr,vr) graph analysis, different and contradicting results were obtained. The slopes of the regression lines in F1 (2004) and F2 were significantly different from zero and unity indicating the presence of gene interaction in the material used. The interception points were below the origin in the three graphs indicating over dominance in the two generations. The array points tacked nearly constant orders between graphs. 4- By the combining ability analysis, seed hardness showed significant specific combining ability (sca) in both evaluation seasons and general combining ability (gca) in the first evaluation season only. The contribution of gca and sca for both seed coat weight and 100-seed weight were significant indicating that both additive and non-additive gene actions determine the expression of these traits. The non-additive genetic variance (sca) was found to be more important than the additive genetic variance (gca) for number of pods/plant and number of seeds/plant. For seed yield, both components of genetic variance were nonsignificant. The parents C.28 and C.86/1 gave the desirable gca effect for all studied traits. Thus they were the good general combiners for these traits in this research. On the other hand, the parent showed undesirable effects for all studied characters. Bulk/1/1/2 Most of the crosses exhibited nonsignificant sca effects for all traits. But, the crosses C.28 × C.86/1 and ZBF/1/1 × C.28 had desirable sca effect for all characters. 5- Yield/plot exhibited significant associations with number of pods/plant, number of seeds/plant and 100-seed weight. High significant and positive associations were observed between number pods/plant and number of seeds/plant. Seed hardness showed positive and significant correlation with seed coat weight and nonsignificant associations with yield and its components in the two seasons. 6- The effect of pods location on the stalk of the plant on the seed hardness and seed coat weight was studied. The pod position had a significant effect on seed hardness in the two seasons. The lower pods showed the greater grand mean for hard seed percentage in the two seasons. Neither the pods position, nor the interactions had significant effect on seed coat weight. 7- Also the seed coat structure of faba bean seed was studied. One line was selected from the high hard seeded group, C.36, and another one from the low hard seeded group, C.28. Transverse sections were prepared from a soft and hard seeds of each line, at the micropyle region and regions away from the micropyle . Photomicrographs were taken from these two regions measurements (in microns) were recorded about the and seed coat thickness, palisade layer, hourglass cells layer, the micropyle and the trachids for all four groups. The main difference between the two lines and between the soft and hard seeds of each line was in the size of micropyle and trachids, which were responsible for water intery to the seeds. The hard seeds had smaller micropyle opening and smaller trachids than soft seeds. Conclusion: Although, seed hardness appeared as a dominant character in the genetic analysis, the parents tacked different orders between the (wr,vr) graphs, indicating fluctuation of the dominance in the two growing seasons and in the two generations. Also indicate the important effect of the environment on this character. The pod position on the stalk also had significant effect on the high percentage of hard seeds. Therefore, the effect of long exposure to temperature, the variability of maturity period, the extend of flowering period and their relation with seed hardness should be studied. In seed coat weight, the parents tacked nearly constant orders between the (wr,vr) graphs, but the gene association had more important role in the degree of dominance, and the pod position on the stalk had no effect. Hence, selection for this character seemed to be more effective and care should be taken since both additive and non-additive factors are included. CHAPTER ONE INTRODUCTION Faba bean is known, in the English Language, by many names, e.g. broad bean, horse bean, field bean, faba bean,….., but the name faba bean has recently been widely used. It is a diploid species (2n = 2x = 12), belonging to the family Papilionasae and is believed to have originated in South West Asia. Botanically, it is divided into three types according to seed size, namely, major (large flattened seed), equinia (medium – sized seed) and minor (rounded small seed). The main cultivars grown in the Sudan belong to the latter two type, but predominantly the medium. Faba bean is the main pulse crop in the Sudan in terms of both area and production, being the main staple food and the main source of protein for millions of people. Its production and consumption have been steadily rising during the last four decades (1960 – 2000). Area and production are shown in appendix (13) for three decades. The area planted to faba bean during the seasons 1999/2000, 2000/2001 and 2001/2002 were 58, 59 and 59 thousand hectars (1 hectar = 2.38 feddans) producing nearly 146 thousand metric tones, each season (FAO, 2002). However, the productivity of this crop is low compared to other crops. Many constraints contributing to this low productivity have been reported by Salih and Salih (1985). Breeding work in faba bean had started at Hudieba Research Station since the early 1960′s and from 1980, the work was also initiated at Shambat Research Station (Salih, 1981, 1983a, Salih and Salih, 1985). The main objectives of the breeding programme were to develop high yielding and stable varieties, with good quality characters and resistance to local diseases. The breeding research programme was achieved through germplasm introduction, hybridization, selection and to a lesser extent mutation breeding. Breeding for seed quality includes breeding for light seed colour and soft seed traits. The local faba bean land races have narrow genetic base, therefore, several hundred accessions were imported from different sources including Egypt, ICARDA, USSR, Ethiopia and Europe, and evaluated. The bulk of the introduced material was not adapted, except those with small seeds. However, the breeding work had resulted in the release Hudeiba 72, SM-L, of many cultivars e.g. BF2/2, Shambat 75, Shambat 104, Shambat 616, Basabeer and Hudieba 93 (Salih and Salih, 1985, 1996; Salih, 1995). Faba bean are characterized by having high percentages of hard seeds and a tough seed coat. Furthermore, most commercial cultivars have not been favorably appraised by consumers tastes, because seeds imbibed water too slowly, which detracts their seed coat to quickly soften and easily be removed during meal preparation. Therefore, recent breeding efforts have been directed towards the production of high yielding stable cultivars with improved quality especially with regard to water imbibing capability and cooking quality. Information on the genetic control of seed hardness, seed coat weight, seed coat thickness and their association with yield and its components appears to be essential for such improvement. The objectives of the present research study were to: 1. study the magnitude of gene action controlling the inheritance of seed hardness and seed coat weight among F1 and F2 generations and their respective parents, as an initial step for making selection to improve these characters, 2. estimate the general and specific combing abilities of parents for seed hardness, seed coat weight, yield and some of yield the components, 3. study the association between these two characters (seed hardness and seed coat weight) and seed yield and its components, and to, 4. study the structural layers of the seed coat to determine their role in water imbibitions. CHAPTER TWO LITERATURE REVIEW 2.1 Seed Hardness Seed hardness is a type of seed dormancy resulting from the impermeability of the seed coat to water or gases from physical resistant to embryo expansion (Salih, 1982 b). In faba bean the occurrence of highly impermeable seed coat is one of the most important factors in delaying germination, affecting cooking quality and reducing the market price (El-Bagoury; 1975; Salih and Ali , 1986). The hard seed character in legume seeds is thought to be due to the thickened palisade layer which is almost impermeable to water and resist the imbibition of it (El-Bagoury, 1975; Esau, 1977; Jha and Singha, 1989). Jha and Singha (1989) found that small incision on the seed coat was most effective in breaking the hard seededness. No single specific cause of hard seed was reported in the literature but, genetic, environmental and crop husbandry factors are thought to be responsible for this phenomenon. Regarding environmental factors, long growing periods resulted in a higher proportion of hard seed (Quinlivan, 1965). Similar El-Bagoury (1975) in increased markedly at maturity in subterranean clover finding was reported in faba bean by Egypt : in that, the hard seed percentage with delaying maturity. This might be attributed to the increase in the hardness of the seed coat tissue as a result of drying and fluctuation of humidity and temperature during this period and their effect on the permeability of the seed coat (Quinlivan, 1965, 1968) and Baciu- Miclaus (1970) showed that the incidence of hard seed was correlated with air humidity. On the contrary, Salih (1982b) found that shortening the growing season increased the amount of hard seed in the cultivar H-72. According to Zenari (1929 ), hard seed character in various Leguminous , Malvaceous and Cistaeous was due to the degree of maturation and therefore can not be considered hereditary. Aitken (1939) demonstrated that the continuity of an impermeable suberized thickening of the top of the malpighian cell, caused hard seed of (Trifolium subterraneum L), and the condition subterranean clover of this thickening depends on genotype environmental conditions and the subsequent degree of dehydration of the seed. High temperature associated with low humidity resulted in maximum hard seededness . Research in the Sudan pointed that, a number of crop husbandry factors affect the proportion of hard seed e.g. sowing date, fertilizer, irrigation and time of harvest (Ayoub, 1971, 1973; Salih, 1978a, 1978 b, 1983 b; Salih and Ali, 1986; Ageeb, 1980; El- Mubarak et al., 1988). Delaying sowing date increases the percentage of hard seed. Salih (1979 a) reported a range of 3.73 to 7.97% of hard 24 seed from respectively. including mid october Ayoub (1971) up showed to November that fertilizer sowing, treatments, nitrogen, phosphorous, potassium, gypsum and animal manure, had an effect on the percentage of hard seed, ranging between 15.0 and 23.0%. Salih (1979 b) found that different levels of nitrogen and phosphorus had no significant effect, but different methods of applying the fertilizers, broad cast, band and placement with seeds had significantly affected the percentage of hard seed. In another experiment, Salih and Ali (1986) showed that the hard seed percentage had increased significantly with increasing levels of phosphorus. Salih (1978 a) reported irrigation that, at Hudeiba, early termination of increased the percentage of hard seed and that by enforcing earlier maturity. On the other hand, Ayoub (1973), at Hudeiba working with BF2/2 increasing watering intervals and Selaim varieties, found that from 8 to 23 days had very little effect on the hard seed percentage. In addition to the above mentioned husbandry practices, the percentage of hard seed was found to be affected by the time of harvest. According to Ageeb (1980), the lowest percentage of hard seed was obtained when the crop was harvested after full maturity. Similarly, Salih (1983 b); Salih and Ali (1986) and ElMubarak et al. (1988) found decreased with that the hard seed percentages delaying harvesting from 90 to 120 days from sowing. Also diseases were considered to be a factor in increasing the percentage of hard seed (Saeed et al., 1987). Considerable variation in the percentage of hard seed among faba bean cultivars was reported in the Sudan. Salih (1976 ) reported a range of 0.1 to 22.8%, and eleven years later, he (Salih , 1987) recorded a range of 1.4 to 13.5%. Smaller range of 5.1 to 11.6% was also reported by Salih and Khairi (1990). Cultivars differ in their hard seed percentage from location to another. In a trial conducted at four locations, namely Shambat, Wad Medani, El-Rahad and New Halfa, Salih and Khairi (1990) recorded a range of 4.7 at New Halfa to 15.1% at Shambat. The cultivar H-72 produced 19.0, 7.8, and 4.4% and the breeding line 00104 produced 10.8, 7.6 and 5.8% of hard seed at Shambat, Wad Medani and New Halfa, respectively. 2.2 Seed Coat Weight The seed coat provides a considerable protection for the enclosed cotyledons and the embryo (Mc Ewen et al., 1974). Faba bean has a tough seed coat which is responsible for the higher fibre content of the seed. This crude fibre detracts from the value of faba beans as a feed and food (Rowland, 1970; Waly and Abdel Aal, 1987). The seed coat accounts about 15% of the weight of the bean seed (Mc Ewen et al., 1974). However, Cerning et al. (1975) revealed that, it constitute a 13.17% of the seed dry weight and it contains 89% of the seed protein. A large variability in the seed coat percentage was reported by many workers ( e.g. Marcellos, 1987; Shehata et al., 1987; Salih and Khairi, 1990 ). Marcellos (1987) examined 30 inbred lines from different sources, viz Mediterranean, Middle East, North Africa and India, for seed coat percentage. He reported a range of 12.2 to 18.2% in a cross locations and also found that the variance fraction for the effect of genotypes was highly significant. Salih and Khairi (1990) reported a range of 14.2% in line 00198 to 16.6% in H-72. Beside that, they studied the variability in testa fraction of ten genotypes at four locations, namely Wad Medani, New Halfa, El Rahad and Shambat, and recorded 17.8, 17.5, 14.2 and 12.1%, for the four sites, respectively. Shaudary and Buth (1970) examined the seed coats of 14 Indian pulses and found that, Vicia faba had the largest palisde and hourglass cells. These cells are primarily responsible for the thickness of the seed coat. Mc Ewen et al. (1974), reported a wide range of variation in thickness of hourglass cells of faba bean. A significant variation in the seed coat thickness was detected by Rowland and Fowler (1977) in faba bean and that due to cultivars and locations. Also, they revealed that the position of the seed on the plant had no effect on the seed coat thickness. 2.3 Inheritance of Seed Hardness and Seed Coat Weight 2.3.1 Inheritance of Seed Hardness The seed hardness character in legumes was reported by many workers (e.g. Altiken, 1939; Forbes and Wells, 1968; Donnelly et al., 1972), to be controlled by both environmental and genetic factors. However, Zenari (1929), reported that, hard seed in various families like leguminasae, Malvaceae and Cistaceae was due to the degree of maturation and could not be considered hereditary. Lute (1928) working with alfalfa (Medicago sativa L.) seed postulated that genetic differences may be obscured by climatic factors. Little work on the inheritance of seed hardness in Vicia faba was reported in the world and no work was conducted in the Sudan. All studies on inheritance of hard seed revealed that, hard seed is dominant over soft seed, and only few genes are involved (e.g. Dhirendra and Singh, 1984; Ramsay, 1997, both working in faba bean; Forbes and Well,1968, in blue lupin; Donnely et al., 1972, in the genus Vicia). In a crossing research work of six genotypes of Vicia faba., Dhirendra and Singh (1984), concluded that dormancy is controlled by both physical and genetic factors. Also dormancy is dominant over non-dormancy and governed by a single gene. Similarly, Ramsay (1997), showed that the segregation patterns for seed dormancy in faba bean is monogenic. This gene, was named doz, and was linked to a gene controlling anthocyanin and proanthocyanidin synthesis. The result of four crosses in the genus Vicia, let Donnelly et al. (1972), to assume a two – gene inheritance for hard seed. Gene A acts as simple dominant for hard seededness. Gene B is dominant for soft seededness when the A locus is homozygous recessive (aa). The double recessive genotype (aabb) is hard seeded. However, they reported some exceptions in F2 as a double recessive epistasis (7 : 9). Lebedeff (1947), in Phaseolus vulgaris L reported that, only a few genes are involved in the differentiation of hard and soft – seeded selections. Bennett (1959) found a rapid increase of hard seed percentage in successive generations of crimson clover suggesting a good heritability of the character. Similarly, Donnelly (1971), when crossed Vicia sativa (2n = 12) with Vicia cordata (2n = 10 ) found that selection for hard seed resulted in an increase in frequancy of plants that produced 70 to 100% hard seed. 2.3.2 Inheritance of Seed Coat Weight Most studies regarding the seed coat characteristics in legumes were concerned with the seed coat thichness (e.g. Rajendra et al, 1979; Ojomo, 1972; Rowland and Fowler, 1977; El Shazly et al., 1995) and colour ( e.g. Bassett, 1997, 1998; Klasinsko, 2000 ). Little studies were converted towards the inheritance of seed coat weight. Through the diallel crossing system, Waly and Abdel Aal (1987), studied the inheritance of seed coat weight by using five parents, and they revealed that the character was under the control of both additive and non – additive gene action. Similar result was reported by El Shazly et al. (1995) and also they concluded that, care showed be taken when planning a breeding programme for selection of thin seed coat, since the genetic behavior of this trait is related to the use of specific genetic material. However, in cow peas, Ojomo (1972) found that the inheritance of seed coat thickness was controlled by two major gene pairs, with a segregating ratios of 9:6:1 and suggesting a duplicate gene interaction. 2.4 Phenotypic and Genotypic Correlations Phenotypic and genotypic correlations between characters give an indication of the characters which may be used as indicators in selection of desired traits (Johnson et al., 1955). The genotypic correlations among characters for which selection is practiced may have important implications in breeding procedures and programmes. Usually the main objectives of the breeding programmes are to develop high yielding and stable varieties with good quality characters. It is known that yield has low heritability because of the strong influence of environment on it. Therefore, determination of the interrelationships with yield structure of a crop and its its components and seed quality traits are essential. Kambal (1969) reported that number of pods per plant had the highest correlation with yield in faba bean. Results obtained by Abdalla (1976) and Magyarosi and Sjodin (1976) confirmed this findng. In contrast Picard and Berthelem (1980) reported that seed weight was the most valuable yield component which can be used as a selection criterion for seed yield potential and yield stability in faba bean. Number of pods per plant is greatly dependent upon the magnitude of the number of podded nodes per plant (Magyarosi and Sjodin, 1976). Strong positive correlations exist between seed yield and number of pods per plant (e.g. Kambal, 1969; Neal and McVetty, 1984; Bakheit and Mahdy, 1988). On the other hand, many workers pointed that number of pods per plant was negatively correlated with number of seeds per pod and seed weight (Mutwakil, 1965; Yassin, 1973; Abu El-Zahab et al., 1980). Number of seeds per plant is an important character in determining grain yield in faba bean and was found to be positively correlated with number of pods per plant (Habetinek et al., 1983). Shalaby and Katta (1976); Mahmoud et al. (1978); Naidu et al. (1985); and Bakheit and Mahdy (1988) found a positive correlation between number of branches and seed yield per plant. Positive significant correlations of number of branches with seed yield, number of pods per plant and number of seeds per plant were obtained by Kambal (1969); Mahmoud et al. (1978) and Sindhu et al. (1985). Seed weight is one of the important yield components in faba bean. Magyarosi and Sjodin (1976) ranked it as a third yield contributing character, following number of pods per plant and number of seeds per pod. Magyarosi and Sjodin (1976); Habetinck et al.(1983); Neal and McVetty (1984) and Hussain et al. (1988) reported a positive relationship between yield and seed weight. 100 seed weight was found to be negatively correlated with number of pods per plant and with seeds per pod (Kambal, 1969; Mahmoud et.al., 1978; Neal and McVetty, 1984). Also negative genotypic and phenotypic correlations of seed weight with number of pods per plant were reported by Yassin (1973). Regarding the seed quality character, Salih (1976), Ramsay (1997) reported a negative association between the hard seed percentage and seed weight. Also Salih (1976) and Rowland and Fowler (1977) found that the final seed yield is not associated with seed coat thickness. In the same experiment, Rowland and Fowler (1977) found that the seed weight and seed coat thickness were not correlated varieties. within varieties but were Waly and Abdel Aal (1987) highly reported correlated a positive among non- significant correlation between seed coat weight and seed weight in both the parents and F1 hybrids. Also, Ikram (1993) found that the seed quality characters exhibited non-significant correlations with yield and yield related characters. Moreover, the correlations between the seed quality characters themselves, were also non- significant. In contrast, Rowland (1970) found that, 1000-seed weight was negatively correlated with crude fiber and positively correlated with seed coat thickness. Also, Marcellos (1987) found that testa fraction increased as seed weight decreased and that was within the tested 30 genotypes of diverse origins. 2.5 Combining Ability The value of an A- or B- line for hybrid production depends on the characteristics of the line as such, and its performance (Kambal, 1962). This latter trait has been called combining ability and is usually subdivided in plant breeding into two categories, general and specific. Spraque and Tatum (1942) defined general combining ability as "the average performance of a line in hybrid combinations". Specific combining ability is used to show those lines in which certain combinations do relatively better or worse than would be expected, on the bases of the average performance of the line involved. General combining ability is associated with genes which are additive in their effects. Specific combining ability, on the other hand, is usually attributed to non-additive types of genes dominance and/or epistasis (Kambal, 1962). such as Bond (1967) and Mahmoud and El-Ayoubi (1986) showed that, selection of an inbred parent for F1 hybrid varieties of faba bean can be made on the bases of its general and specific combining abilities. Most of the studies in faba bean revealed a significant combining abilities for seed yield and its components and seed quality traits (e.g. Kaul and Vaid, 1996; El-Hosary et.al., 1997; Waly and AbdelAal, 1987; El-Hosary, 1984, 1985). However, the seed yield and its components found more interest than seed quality traits in research. 2.6 Seed Coat Structure and its Anatomy Faba bean has been characterized as having a tough seed coat which causes difficulties in grinding and is responsible for seed high in fibre content (Rowland and Fowler, 1977). Furthermore, Bell (1975) and Rowland (1970) postulated that, seeds of faba bean have much higher crude fibre level than the seeds of many other legumes. This crude fibre is located mainly in the seed coat and detracts from the use of faba beans as feed and food. Esau (1979) reported that the presence of palisade layer in seed coats of certain hard legume seeds is assumed to be causally connected with their degree of impermeability. Before that, Shaudaey and Buth (1970) examined the seed coats of 14 Indian pulses and found that, the Vicia faba had the largest palisade and hourglass cells. These cells are primarily responsible for the thickness of the seed coat. A wide range of variation in the thickness of hourglass cells and palisade of faba bean had bean observed by Mc Ewen et.al. (1974) and. Youssef and Bushuk (1984) Mc Ewen et.al.(1974) determined the structure of seed coat of faba bean variety Akerprle using scanning electron microscope. The photomicrographs showed no discontinuity in the thick seed coat. Cross section of the seed coat showed characteristic palisade, parenchyma, trachid and hourglass cell. They showed a full description of faba bean seed layers. And they indicated that: The hilum or seed scar, play an active role in the dehydration process of the bean. Tissues on the surface of the hilum appears flaky and fold. A cross-section of the seed coat in the hilum region showed several distinct types of tissues. Immediately under the flaky surface layer of the hilum there is a double layer of palisade cells, where as the inner layer appears to be continuous with the single palisade layer found throughout the seed coat. The trachid cells are below the central groove of the hilum. Arround the trachid cells there are loosely and irregularly structured parenchyma cells. In regions away from the hilum, between the palisade and the parenchyma cells , the hourglass cells are located, which show a variety of shapes and sizes. Youssef and Bushuk (1984) demonstrated that the micropyle is situated just below the hilum in legume seed. Also they explained that the hard - to cook Egyptian types, their seeds had smaller micropyle opennings, shorter hourglass cells and thicker cell layer than those of soft - to cook types. This micropyle governs water entry to the cotyledon as shown by Powerie et.al. (1960) and Yossef et.al. (1982). CHAPTER FOUR RESULTS 4.1 Inheritance of Seed Hardness and Seed Coat Weight 4.1.1 Inheritance of Seed Hardness 4.1.1.a Using Genetic parameters Analysis of variance according to Hayman(1954), (Table 2) and the estimation of components of genetic variation (Table 3) revealed that most variations in seed hardness were attributed to non-additive gene effect. In that, the items b, b2 and b3 (non-additive factors) were significant in both F1 and F2. However, the item b1 showed nonsignificant effect in the two generations. The non-additive genetic components of variations, H1 and H2, were significant in both F1 and F2, while the additive genetic component, D, revealed nonsignificant effect (Table 3). The estimates of F (average covariation of additive and dominance effect over all arrays) had positive nonsignificant values in the two generations. The estimates of h2 (weighted average dominance factor effect) were positive in F1 and negative in F2. The environmental variance, E, was also significant in the two generations. Regarding the genetic parameters ratios (Table 3), the estimates of ½ average degree of dominance, (H1/D) , were 1.5, 3.9 and 2.28 for the F1 in the first evaluation season (2003), F1 in the second evaluation season (2004) and F2, respectively. Also, the average values for uv (frequencies of positive and negative alleles ) over all loci were 0.15, 0.21 and 0.2, respectively, as presented by the ratio H2/4H1 (Table 3). Table 2: Analysis of variance and mean square Of genetic variances for seed hardness in of faba bean (V. faba L) in the two seasons, 2002/3-2003/4. Sourse of variation d.f F1(2003) F1(2004) F2 Treatments 35 22.52** 39.84** 36.75** Replications 1 38.57** 3.45 30.36** Error 35 5.83 17.14 11.28 LSD 4.90 8.4 6.72 C.V.% 36.0 35.1 26.7 S.E 2.42 4.14 3.36 Additive variance a 7 58.067 67.46 42.730 b1 1 20.927 64.68 1.19 b2 7 92.639* 57.37* 33.67* b3 20 24.880** 83.95** 66.02** b 28 41.679* 76.61** 55.61** total 35 78.30 136.07 97.53 a × blocks 7 24.73 23.16 16.81 b1 × blocks 1 10.67 6.57 13.09 b2 × blocks 7 25.23 12.62 8.39 b3 × blocks 20 5.44 18.97 18.13 b × blocks 28 14.58 16.47 14.58 total × blocks 35 28.27 30.98 26.69 non-additive variance *p ≤ 0.05, **p ≤ 0.01. Table 3: Components of genetic variance, their standard errors and the estimated genetic parameters for seed hardness in faba bean (Vicia faba L.), in the two seasons, 2002/3-2003/4. Genetic variance F1 (2003) F1 (2004) F2 additive genetic variance D 18.91 ±10.56 4.17± 2.89 6.64±4.49 45.11*±10.53 25.92*± 9.16 1.1* ± 0 .179 32.91 ± 8.82 3.37* ± 1.53 63.71*±0.78 53.62*± .91 2.55 ± 7.38 7.56 ± 0.61 8.38*± .32 138.09*± 2.65 128.34*±7.44 -81.73*±5.11 16.26± 2.07 5.91*±1.56 1.5 0.15 3.9 0.21 2.28 0.20 3.2 -0.5* 0.25 0.05 20.9 1.6 -.35* 0.123 0.05 13.1 3.12 -0.34* 0.120 -0.64 11.7 non-additive genetic variance H1 H2 h2 F E genetic ! parameters ( H1/D)½ H2/4H1 [(4DH1 ) ½ + F]/ [(4DH1 ) ½ - F] r r2 h2/H2 Heritability (%) *p ≤ 0.05, **p ≤ 0.01. for genetic parameters ratios in F2 see material an methods pp 23 ! The proportion of dominant and recessive genes [(4DH1))½+F]/[(4DH1)½–F], approximately, ranging between two to three genes and the ratio h2/H2 (number of genes which control the character and exhibit dominance ) gave 0.05 and 0.64 values in the two generations, respectively. The coefficient of correlation (r) between the parental order of dominance (wri + vri) and the parental measurement (yri) gave negative and significant values in the two generations (Table 3). Heritability percentages were 20.9, 13.1 and 11.7 in F1 (2003), F1 (2004) and F2, respectively. 4.1.1.b using (wr,vr) Graph To evaluate the inheritance of seed hardness of faba bean by means of (wr,vr) graph analysis, the wr (covariance of arrays ) values were ploted against the corresponding values of vr (variance of arrays ), (appendix 3 for F1(2003), F1(2004) and F2) . From all, the three graphs (figures 1, 2 and 3), the following results could be readied. The general picture of the graphs appeared to be similar. The array point exhibited considerable scatter around the regression lines in each graph, the regression line was not significantly different from zero or unity i.e. b = 0.55 ± 0.45, 0.48 ± 0.52 and 0.404 ± 0.33 in the three graphs, respectively. Also each line intercept its wr axis a short distance below the origin, in that, the interception point, a, had negative sign ( a = -3.66, -8.26 and -3.69), respectively. However, the array point orders were different between graphs. In F1 (2003) (Fig 1), the array points 2(C.28) and 4(C.36/1) are near to the origin (P > 75% dominant genes), array 5(ZBF/1/1), 8(C.36), 3(C86/1), and 1(BB7) were middle 7(C.22), in their positions (50 < P < 75%). On the other hand the point for array 6(Bulk/1/1/2) is far (away) from the origin. In F1 2004 the array points 1, 3, 5 and 6 were near to the origin, while 4, 7 and 8 in the middle and 2 was far away from the origin. Regarding the F2 data, the array point 1 was near to the origin. The points of the arrays 5, 6, 2, 3 and 7 were middle in their position and also the point for array 3 was far away from the origin. 4.1.2 Inheritance of Seed Coat Weight 4.1.2.a using genetic parameters Hayman (1954) analysis of variance is presented in Table 4 and Table 5 showed the estimates of components of genetic variance in seed coat weight. Both additive (a) and non-additive (b) gene effects were significant in F1 and F2. The items b and b3 had significant effects in the two generations, while b1 showed non signification effect, and b2 had significant effect in F1 only (Table 4). The estimates of genetic variance component (Table 5), confirm the results of the means of squares. The statistics representing both, the additive (D ) and non-additive (H2 and H1) gene effects were significant in the two generations. The average dominance (h2) showed negative values in the two generations, and the F estimates had significant and positive values. The environmental variance (E) was also significant in the two generations. The genetic parameter ratio (H1/D)½ was 1.5 in F1 (2003), 0.57 in F1 ( 2004) and 0.95 in the F2. The gene frequency, uv, ratios were 0.20, 0.19 and 0.12, respectively. Proportion of dominant and recessive genes were three to four genes and the average dominant gene ratios (h2/H2) were -0.21, -0.2 and -0.35 values in F1 (2003), F1 (2004) and F2, respectively. The coefficients of correlation (r) between parental order of dominance (wri+vri ) and the parental measurement (yri) gave negative values in the two generations. The values of heritability estimates were low to moderate as follows 13.7, 51.3 and 20.8% in F1 (2003), F1 (2004) and F2, respectively. 4.1.2.b using (wr ,vr ) Graph figures 4, 5 and 6 showed the (wr ,vr ) graphs for seed coat weight in F1 (2003), F1 (2004) and F2 , respectively. The regression lines slopes were 0.69 ± 0.42, 1.65 ± 0.45 ± 0.31 in F1 (2003), F1 (2004) and F2, respectively. and 1.69 The interception points showed negative values, -1.14, -0.57 and -2.13, respectively. However, graphs the array points tacked nearly constant orders between wr 25 20 15 6 10 4 5 0 -5 7 1 vr 3 2 -5 0 5 10 8 15 20 25 5 -10 -15 -20 -25 Fig 1: ( wr-vr) graph for hard seed of faba bean in F1 ( Season 2002/2003) b = 0.55 ± 0.45 a = -3.66 — w –⋅–⋅ . w rei ri (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays wr 40 30 20 10 2 6 1 0 -10 7 3 4 5 0 10 20 30 8 40 50 60 -10 -20 -30 Fig 2 : (wr,vr )graph for hard seed of faba bean in F1( season2003/2004) b = 0.48 ± 0.52 a = -8.26 — w –⋅–⋅ . w rei ri (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays 7 wr 30 25 20 15 10 2 5 1 5 6 0 -10 7 -5 0 3 10 204 v 30 8 -10 -15 -20 Fig 3: ( wr, vr) graph for hard seed of faba bean in F2 b = 0.404 ± 0.33± a = -3.69 — w –⋅–⋅ . w rei ri (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays 40 Table 4: analysis of variance and mean squares of genetic variances for seed coat weight in faba bean (Vicia faba L) in the two seasons, 2002/3 - 2003/4. Source of variation Treatments Replications Error LSD C.V% S.E additive variance a non-additive variance b1 b2 b3 b total a × blocks b1 × blocks b2 × blocks b3 × blocks b × blocks total × blocks *p ≤ , **p ≤ 0.01 d.f 35 1 35 F1(2003) 3.76** 6.07** 1.23 2.25 8.9 1.11 F1(2004) 3.36** 0.52 0.97 2.003 7.2 0.99 F2 2.46** 0.03 0.58 1.52 5.95 0.76 7 11.602* 23.35* 8.89* 1 7 20 28 35 7 1 7 20 28 35 1.43 4.897* 7.83* 6.87* 13.31 3.36 1.82 1.46 2.67 2.21 4.6 0.097 2.93* 2.1** 2.24** 8.26 5.53 0.315 0.54 0.14 0.024 2.14 3.35 10.83 1.47** 3.87* 7.98 2.64 0.36 5.08 0.15 1.39 2.98 Table 5: Components of genetic variance, their standard errors and the estimated genetic parameters for seed coat weight in faba bean (Vicia faba L.), in the two seasons, 2002/3-2003/4. Genetic F1(2003) variance additive genetic variance D 2.48*±0.92 non- additive genetic variance H1 5.61*±2.13 H2 4.91*±1.85 h2 -1.01* 0.24 F 2.60 ±2.20 E 0.679* ± .31 genetic ! parameters ( H1/D)½ H2/4H1 [(4DH1 ) ½ + F]/ [(4DH1 ) ½ - F] r r2 h2/H2 Heritability (%) F1(2004) F2 4.04*± 0.32 4.44* ± 0.31 1.32* ± 0.67 0.999* ± .52 -0.198 0.43 2.80* ± 0.76 0.48* ± 0.11 16.35* ± 2.81 7.31* ± 2.44 -2.61* ± 0.64 12.80* ± 1.44 0.27* ± 0.10 1.5 0.2 0.57 0.19 0.95 0.12 3.6 -0.5* 0.25 -0.21 13.7 4.01 -0.8** 0.64 -0.2 51.3 4.09 -0.6** 0.36 -0.35 20.8 *p ≤ 0.05, **p ≤ for genetic parameters in F2 see pp 23 ! wr 5 4 3 2 2 1 3 7 1 8 5 0 vr 6 -10 -5 -1 0 5 10 15 4 -2 -3 Fig 4: ( wr,vr) grahp for Seed coat weight of faba bean in F1 (season 22/2003) b = 0.69 ± 0.42 a = -1.14 — w –⋅–⋅ . wri rei (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays wr 6 4 1 3 2 4 0 -1 5 6 7 2 8 vr 0 1 2 3 -2 -4 -6 seed coat weight of faba bean in F1 ( Season Fig 5: (wr,vr) graph for 2003/2004) b = 1.65 ± 0.45 a = -0.57 — w –⋅–⋅ . w rei ri (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays wr 8 6 4 1 2 6 7 0 -1 -0.5 0 4 0.5 3 8 5 2 vr 1 1.5 2 2.5 3 -2 -4 -6 Fig 6: (wr,vr) graph for seed coat weight of faba bean in F2 b = 1.69±0.31 a = -2.13 — w –⋅–⋅ . w rei ri (expected regression line of b = 1 ) wreii (estimated regression line) points orders of the arrays 3.5 4.2 Combining Ability The investigated genotypes, F1 and their respective parents, showed highly significant differences (Table 6) for seed hardness and seed coat weight in the two evaluation seasons ( 2002/2003 and 2003/2004 ). However, significant differences were observed for seed yield /plot, number of pods/plant, number of seeds/plant and 100-seed weight in the second season only. Combining ability analysis was restricted only in F1 towards characters in which genotypes exhibited significant difference Means squares for combing ability for all characters are summarized in Table 7. Seed hardness exhibited significant specific combing ability ( sca ) in both seasons and significant general combing ability ( gca) in the first season only. Moreover, signification gca and sca were observed in seed coat weight and 100-seed weight in both seasons, but number of pods/plant and number of seeds/plant gave only significant sca. However, seed yield/plot had nonsignificant combing ability. Estimates of gca effects for individual parents for all traits from the F1 generation are presented in Table 8. The parental lines 2(C.28), 3(C.86/1) and 4( C.36/1) gave significant positive gca effect for number of pods/plant, number of seeds/plant and 100-seed weight. Moreover, they showed negative significant effect in seed hardness and seed coat weight, except the showed negative significant and 8(C.36) gave 4(C36/1) which effect for seed coat weight in the second season. On the other hand, 7(C.22) parental line the parental lines 5(ZBF/1/1) significant gca effect for high percentages of hard seed and seed coat weight in both seasons. Line 1(BB7) had the highest gca effect for lower seed coat weight percentages in the two seasons, but it gave negative values for number of pods/plant and number of seed/plant. Furthermore it gave positive effect for high percentages of hard seed in the second season. The parental line 6( Bulk/1/1/2) showed undesirable effects for all characters studied . Table No. 9 spresent the estimates of sca effects of individual cross combinations for all characters studied in F1. Most of the crosses exhibited nonsignificant sca effect for all traits. For seed hardness the crosses 1×3, 1×7, 2×5, 2×6, 3×5, 5×6, and 7×8 gave high positive and significant sca effects. The crosses 2×3, 2×7, 5×8, and 6×8, showed negative significant sca effects. However the crosses 1×6, 3×7 and 5×7 had negative and significant effects in the first season only. Regarding seed coat weight, most of the crosses showed nonsignificant sca effects. The highest negative values were given by cross 4×7 followed by cross 1×3 in both seasons . However, crosses 1×6 and 2×6 gave the highest positive effects. The crosses 2×3, 2×4, 2×5, 2×6, 3×6, 4×5, 4×6, 4×8, 5×6, and 5×8 showed significant and positive effects for two or three traits of yield and its components. Table 6: characters Source of d.f vriation Seed hardness I Treatments 35 22.53** Replications 1 38.57 error 35 5.833 Lsd C.V% S.E. Mean squares from analysis of variance for all characters studied in faba bean (Vicia faba L) in the two seasons, 2002/203-2003/2004 4.903 36.0 2.42 II 39.84** 3.45 17.14 8.404 35.10 4.14 Seed coat weight I II Pods/plant I 3.76** 3.36** 3.57 6.07 0.517 12.251 1.23 0.973 7.96 2.25 8.9 1.11 2.003 7.2 0.99 5.73 35.2 2.82 Seeds/plant 100-seed weight II I II I II 74.72** 15.49 28.72 30.22 144.22 50.00 282.7** 33.6 131.00 54.17 22.67 42.89 38.77** 56.003 8.206 10.88 19.6 5.36 14.36 35.4 7.07 23.24 28.50 11.4 13.296 4.8 6.9 5.82 5.70 2.87 *p≤ 0.05, **p≤ 0.01. I, II were the seasons 2002/2003 and 2003/2004, respectively. Yield/plot I II 0.0396 0.221** 0.016 0.18 0.086 0.0794 0.597 8.10 1.106 0.573 7.3 0.99 characters Source of variation General combining ability(gca) Specific combining ability(sca) Table 7: Mean squares from combining ability analysis for all characters. (Seed hardness and seed coat weight in the two seasons, and other traits in the last season). Seed hardness Seed coat weight Pods/plant Seeds/plant 100Yield/plot *p≤ seedweight 0.05, F1(2003) F1(2004) F1(2003) F1(2004) **p≤ d.f 0.01. 7 10.53** 17.31 2.66** 5.84** 26.89 108.88 42.59** 0.053 28 11.44** 19.43* 1.69** 1.89* 39.88** 149.15* 13.07** 0.14 105 2.9 8.57 0.62 0.487 14.36 6551 4.10 1.39 Component due to gca 1.16 0.874 0.204 0.535 1.25 4.34 3.85 -0.039 Component due to sca 7.5 10.86 1.07 1.4 25.52 83.64 8.96 -1.25 ratio 0.15 0.08 0.20 0.38 0.05 0.05 0.43 0.03 error Table 8: Estimates of the general combining ability effects from the first generation for all characters in the seasons 2002/2003-2003/2004 Characters Seed coat weight Seed hardness parents BB7 C.28 C.86/1 C.36/1 ZBF/1/1 Bulk/1/1/2 C.22 C.36 S.E(gca) S.E(gi-gi) F1(2003) Pods/plant F1(2004) F1(2003) Seeds/plant F1(2004) 100-seed weight Yield/plot 0.556 - 2.57* - 2.29 - 0.07 0.31 0.224 1.068* 1.369* - 0.204 -1.131* -2.104** -1.104 1.696** 0.004 0.729 2.321** - 0.97** - 0.45* 0.00 - 0.22 0.54* 0.05 0.52* 0.53* -1.124** - 0.659* - 0.624* 0.651* 1.016** 0.301 0.204 0.641* - 0.495 1.82* 1.915* 1.595* - 0.336 -2.395* 1.095 -2.20* -5.861** 2.657* 2.542* 3.759** 1.347 - 4.278** 1.947 0.887 2.246** 1.266* 3.241** -2.109** -1.629* -1.674* 0.291 -1.634* 0.068 0.018 0.069 0.069 -0.096 -0.11 0.018 -0.037 0.51 0.76 0.47 1.31 0.22 0.34 0.21 0.31 1.12 1.7 1.75 3.6 0.47 0.91 0.059 0.089 *p≤ 0.05, **p≤ 0.01 Table 9: Estimates of specific combining ability effects of individual cross Combinations for all characters studied in F1 generation Characters Seed hardness cross 1×2 3 4 5 6 7 8 2×3 4 5 6 7 8 3×4 5 2003 0.094 3.79** -0.21 -1.47 -4.5** 3.80** 4.25** -2.67* 1.27 1.96* 1.95* -2.42* 0.53 1.42 5.86** Seed coat weight 2004 -4.43* 3.8** 1.3 1.00 0.7 4.32* -1.37 -4.78 8.71** 4.42* 14.35** -4.91* 11.04** 0.95 4.4* 2003 -1.31 -1.76 1.86* 1.29 1.89* -0.08 1.106 -0.82 1.24 0.98 10.11** . 039 1.29 0.087 -0.97 2004 -0.316 0.199 0.373 0.409 1.124* 0.179 1.134 -0.716 0.809 0.694 11.18** 0.264 0.769 0.324 0.009 Pods/plant Seeds/plant 2004 -5.28* -2.126 -3.606 -3.426 -1.466 5.64* 1.59 4.66* 5.48* 9.66** 25.5** 3.48 0.37 2.33 6.26* 2004 -8.29 -3.67 -4.27 -4.53 -2.05 9.77 -4.12 4.26 0.34 7.95 6.724 4.504 16.514* 10.557* 7.769 100-seed weight 2004 1.45 2.225 5.325** -0.405 4.39** 2.425 -2.15 6.205** -3.445* 3.075* 2.62* -2.595 -0.67 -1.42 -0.65 Yield/plot 2004 0.109 -0.191 -0.233 0.066 0.021 0.174 0.137 0.036 0.632** -0.344 0.711** 0.078 0.247 0.12 0.315 Table 9: Cont. 6.32* 0.624 1.10 -0.48 2.02* -2.605* 4.49 6 0.354 10.369* 0.03 0.579 0.83 1.125 -1.32 -1.96 7 0.121 3.38 0.584 0.78 0.137 -0.395 -1.621 0.03 8 0.136 * * -0.67 -0.667 -0.60 -0.35 2.95 11.102 0.159 4×5 0.35 ** * 9.49 -0.75 -1.401 1.44 2.695 -5.07 1.23 6 0.334 0.454 -4.17* -3.63* 0.78 0.40 3.60 1.08 7 0.29 * ** ** -1.24 1.28 -1.24 4.455 4.75 18.76 1.58 8 0.416* 0.034 3.8* 2.87 12.39* 3.9858 0.374* -1.023 3.84** 5×6 * -1.51 3.63 1.55 -2.38 0.307 1.03 0.76 -0.08 7 * ** ** ** * -0.41 8.23 2.725 -2.18 -0.403 -7.52 18.52 0.41* 8 0.54 -0.503 2.54 4.29 3.845** -0.185 6 ×7 -1.11 -2.77 ** ** -1.14 -0.313 -2.96 -7.60 -0.32 8 -3.51 -5.32 0.52 ** ** * -0.54 0.017 -3.85 13.73 -0.36 4.01 3.40 7 ×8 2.555 ——————— ————— ——— ———— ————— ————— ————— ———— ———— 0.63 0.7 1.07 1.83 0.181 S.E(sca) 5.1 2.36 1.3 0.76 0.58 1.87 3.2 0.22 S.E(sij-sij) 8.87 4.15 2.22 1,2,3,........,8 are the parents, BB7, C.28, C.86/1, ZBF/1/1, Bulk/1/1/2, C.22 and C.36, respectively. *≤ 0.05; **p≤ 0.1. 4.3 Interrelationships Between the Different Characters Estimates of phenotypic correlation coefficients for various pairs of characters, in all possible combinations, are presented in Tables 10 and 11 for the two evaluation seasons (2002/2003, 2003/2004), respectively. In both seasons, yield/plot showed highly positive significant correlations with number of pods/plant, number of seeds/plant and 100-seed weight. Also highly significant and positive association was observed between number of pods/plant and number of seeds/plant in both seasons. However, 100-seed weight exhibited nonsignificant correlation with these two traits. Seed hardness and seed coat weight exhibited significant and positive correlation ( at 0.05) in both seasons. Seed hardness showed nonsignificant association with yield and it′s components in the two seasons, but at the same time, seed coat weight had significant negative correlation with yield and it′s components in the second season only Table 10: Phenotypic correlations between six pairs of characters in eight faba bean genotypes evaluated in season 2002/203. characters Seed coat weight Seed hardness Seed coat weight Pods/plant Seeds/plant 100-seed weight *p≤ 0.05; **p≤ 0.1. 0.44* Pods/ plant Seeds/ plant 100-seed weight Yield/ plot 0.043 -0.047 0.162 0.004 -0.171 -0.155 - 0.099 0.215 0.923** - 0.204 0.607** - 0.102 0.592** 0.504** Table 11: Phenotypic correlations between six pairs of characters in eight faba bean genotypes evaluated in season 2003/2004 characters Seed hardness Seed coat weight Pods/plant Seeds/plant 100-seed weight *p≤ 0.05; **p≤ 0.1 Seed coat Pods/ weight plant Seeds/ plant 100-seed Yield/ weight plot 0.328* -0.068 -0.363* -0.01 -0.653** -0.679** -0.095 0.91** -0.035 0.851** -0.091 -0.50** -0.383* 0.861** 0.338* 4.4 Effect of Pod Position on Seed Hardness and Seed Coat Weight The effect of pod position on the stalk of the plant on seed hardness and seed coat weight was studied. The results were presented in Table 12 and Appendices 11 and 12 For the two traits the lines exhibited variability in the two seasons. The pod position had significant effect on seed hardness in the two seasons. However, the interaction between the lines and pod position was not significant. The lower pods showed the greater grand mean for hard seed percentages in the two seasons. They gave 8.5 and 9.2% in the first and second season, respectively, while the upper pods had 5.34 and 7.5% in the two seasons, respectively (appendix 11). Regarding interaction the had seed coat, neither significant the effect. pod position, nor the The two pod positions, approximately, showed equal grand means (appendix 12). Table 12: Mean squares for seed hardness and seed coat weight in faba bean, showing the significant effect of pods position on the plant Characters Seed hardness Seed coat weight Source of variation d.f (2003) (2004) (2003) (2004) replication 1 13.78 40.86 0.42 0.085 Parents 7 80.56** 35.46* 11.74** 11.494** Position 1 55.65 30.12* 0.061 0.72 parent×position 7 15.04 7.81 0.224 0.346 error 15 6.11 5.1 0.43 0.499 Lsd(parents) 3.73 5.66 0.98 1.064 Lsd(position) 1.86 2.83 0.49 0.532 S.E 0.93 3.76 0.65 0.706 C.V% 12.3 42.1 5.2 5.4 *p≤ 0.05; **p≤ 0.1. 4.5 Seed Coat Structure Photomicrographs of cross sections of faba bean seed coat were presented in figures 7 and 8. A cross sections of seed coat in region away from the micropyle showed several distinct layers of tissues (Fig 7 a). The outer most layer, the epidermis (cuticle), followed by a single palisade layer of sclereids (macrosclereids), hourglass cells, parenchyma cells and the endosperm. The micropyle opening , which stay at one end of the hilum, appeared in a triangle shape (Fig 8 a). This region consist mainly of a compact group of trachids (Fig 8 d) surrounded by a pocket of parenchyma cells and two palisade layers at the outer surface. Measurements were recorded about seed coat thickness, length and width of palisade cells, hourglass cell, trachids and the micropyle in microns (1 micron =10-6 m) and tabulated separately (Tables 13, 14, 15, and 16). Seed coat thickness: Different values were obtained for seed coat thickness in the four samples studied (the line C.28, soft and hard seeds and line C.36, soft and hard seeds ). In C.28, thickness of the soft seeds was 427.5 µm away from the micropyle and 549 µm near to the micropyle. While the the thickness of the hard seeds was 369 µm and 540 µm in the two regions, respectively. Line C.36, which belongs to the hard seeded group, recorded 405 µm away from the micropyle and 535.5 µm near the micropyle in the soft seeds. The hard seeds had a thickness of 360 µm away from the micropyle and 472.5 µm near the micropyle region. Table (13): seed coat thickness (in microns) Soft seeds Hard seeds Lines ↓ I II I II C.28 427.5 549 369 540 C.36 405 535.5 360 472.5 I: away from the micropyle, II: near the micropyle. Palisade cells: Equal values of 360 µm for width of palisade cells, in line C.36 for soft and hard seeds and also for hard seeds of line C.28. Also, approximately near to equal length in the three samples were measured ,198, 193.5 and 202.5 µm, However, the soft seeds of line C.28, respectively (Table14). measured 45 µ m in width and 225 µm in length Table (14): palisade cells (in microns) Soft seeds Hard seeds Lines ↓ Width length width length C.28 45.0 225 36.0 202.5 C.36 36.0 198 36.0 193.5 Hourglass cells: The hourglass cells showed different sizes and shapes. The width measured a range of 67.5 - 99 µm in all four groups. Also the soft seeds measured a range of 112.5 – 157.5 µm in length. Table (15): Hourglass cells (in microns) Soft Lines ↓ seeds Hard seeds Range of Range of Range of Range width length width length C.28 67.5–99 112.5–157.5 67.5–99 67.5–157.5 C.36 67.5–99 112.5–157.5 67.5–99 67.5–157.5 of Micropyle and trachids: A clear difference in measurements of the micropyle and the trachids between the four groups and between the soft and hard seeds in the same line were observed (Table 16). In line C.28 the micropyle of the soft seeds measured 2070 µm in width and 1540 µm in length, while the hard seeds measured 860 µm in width and 590 µm in length. Line C.36 had 1380 µm width and 1280 µm length for soft seeds. However, the hard seeds gave 820 µm and 450 µm for width and length, respectively. Table (16): micropyle and trachids measurements (in microns) Soft Line ↓ C.28 Width Hard seeds length width length 1541 860 590 989 270 295 micropyle 1380 1280 820 450 trachids 220 300 155 300 micropyle 2070 trachids C.36 seeds 494.5 The trachid cells group in C.28 measured 494.5 µm in width and 989 µm in length in soft seeds and 273 µm in width and 295 µm in length for hard seeds. The soft and hard seeds in C.36 measured equal length of the trachids. However, the two groups got different widths, 220 µm in the soft seeds and 155 µm in the hard seeds. Epiderms (Cuticle) (Palisades) (Hourglass cells) (Parenchyma cells) Hourglass cells (a) Palisade cells (Endosperm) (Cotyledon) (b) Fig (7): photomicrographs of cross section of faba bean seed coat, showing several distinct types of cells. (b) Enlarged view of palisade cells and hourglass cells. (a) (a) (b) (c) (Parenchyma cells) (Palisade layers) (Trachid cells) (d) Fig (8): photomicrographs of cross - section in the micropyle region. (a) The triangle shape of the micropyle, (b) Enlarged view of trachid cells, (c) Enlarge view of parenchyma cells, (d) A compact group of trachids. CHAPTER FIVE DISCUSSION 5.1 Inheritance of Seed Hardness and Seed Coat Weight 5.1.1 Inheritance of Seed Hardness 5.1.1.1 Genetical parameters It was clear from the results of both analysis of variance (Table 2) and the components of genetic variations (Table 3), that seed hardness was a dominant character and the non-additve gene effects(b, H1 and H2) were more important in the inheritance of this character. The nonsignificance of b1 item indicated the ambidirectional dominance of the character. Moreover, the genetic variance item H2 showed significant values in both F1 and F2 indicating a dominance asymmetry (unequal distribution) of positive and negative effects of genes, and the resultant values for the dominant item b2 and H2/4H1 ratios tend to confirm this result. The values of H1/D (mean degree of dominance), provide evidence for the prevalence of over dominance in the inheritance of seed hardness ½ in the experimental material, [( H1/D) ¼(H1/D) > 1 in F2]. ½ > 1 in F1 and Positive F estimates were obtained in the two generations indicating the predominance of the dominant alleles in the parents used, which was in line with the values obtained for r (correlation coefficient between the parental mean, yri, and the parental order of dominance, wri+vri, which showed negative and significant estimates. Singh and Shaudary(1977) postulated that, if the correlation is negative, it means parents containing most increasing genes have the lowest values of (wri+vri), and thus contain most dominant genes and vise versa. Agroup of 2 to 3 genes were observed in the inheritance of seed ½ ½ hardness, in that the ratio [(4DH1)) +F]/[(4DH1) –F] exhibited a range of 2.6 to 3.0 indicating its quantitative inheritance. However, a single dominant gene was observed to control the character (h2/H2, approx. = 1). Many workers reported a single dominant gene in controlling seed hardness as indicated in the literature review. Despite of the dominant effect of the gene, the character had low heritability in this study. This showed that the environmental effect was more important, which was confirmed by the significant values of the environmental components of variance (Table 3). Also, the additive genetic variance estimates, D, were small compared to the non-additive ones (H1 and H2). Falconer (1980) showed that the heritability is a property not only of a character but also of the population and the environmental condition to which the individuals are subjected. Furthermore, Johnson el. al.(1955), in a study of a number of characters in soybean, concluded that estimates of heritability may vary greatly depending upon the character, the population and the sample size. 5.1.1.2 (wr,vr) Graphs The (wr,vr) graph provides tests of significance for the presence of dominance (b ≠ 0) and the average degree of dominance (sign of a), in which b is the slope of the regression line and a is the wr intercept (Hayman, 1954). The (wr,vr) graphs for seed hardness of faba bean (Figures 1, 2 and 3) in F1 and F2 also revealed seed hardness as a dominant character. In the three graphs, the slopes of the regression lines, b, were not significantly different from zero or unity. This suggested the absence of gene interaction in the expression of the character (Hayman, 1954; Whitehouse et. al., 1958; Gomathinayagam and Dabholkar, 1992). In the experimental material used, in the two generations, each regression line intercepted the wr axis below the origin. Thus, the (wr,vr) graphs also indicated that there was an over dominance in the expression of seed hardness. The position of the arrays points along the line of wr on vr depend on the relative proportion of dominant and recessive alleles present in the common parent of each array (Hayman,1954; Jinks,1954). "Parents possessing excess of dominant alleles will have a low array variance and covariance, and will lie near the origin. Highly recessive parents will have a large array variance and covariance, and will lie at the opposite end of the regression line. If the dominance effects of the genes are unequal, the position of an array point will be weighted in favour of genes with large dominant effects (Crumpacker and Allard, 1962)". The eight array points exhibited considerable scatter around the regression lines. This provides evidence for genetic However, the order of these points diversity between them. differ in the three graphs. Array 1(BB7) appeared to posses near to 50% dominant genes in F1(2003), but, in F1(2004) and F2 appeared to have approximately 100% dominant genes. Similarly, parents 2(C.28) and 6(Bulk/1/1/2) changed their orders. Whitehouse et al.(1958) postulated the differences of array orders as a reflection of the different growing conditions. The correlation coefficients (r) of yri and (wri+vri) were found to be negative and significant. This provides evidence that most of the dominant alleles in the parents are acting in the direction of high percentage of hard seed, which was also indicated by the positive values of F estimates. Therefore, the majority of the arrays points on the (wr,vr) graphs lie in the half closer to the dominant end. 5.1.2 Inheritance of Seed Coat Weight 5.1.2.1 Genetical parameters The statistical analysis for seed coat weight revealed that the character was under the control of both additive and non-additive gene action. Waly and Abdel Aal (1987) and El-Shazly et. al.(1995) reported similar results in faba bean. The nonsignificant values of b1 item indicated ambidirectional dominance of the character. The non-addidive genetic component of dominance ,H2, and the mean square of the genetic component b2 were significant in both F1 and F2 indicating an unequal distribution of increasing and decreasing genes in the parents, which was in the same line of the value of the uv ratio (H2/4H1 <0.25), (Table 5). The significant and positive F estimates for seed coat weight in the two generations indicated that, dominant alleles are more frequent than recessive ones. At the same time, the coefficients of correlation (r) tend to confirm this result, where they gave negative significant values. Average degree of dominance involved in the action of genes was in the range of partial dominance in F1(2004) and F2 and over dominance in F1(2003) as indicated by the ratio (H1/D)½. This difference may be due to gene association. Gomathinayagam and Dabholkar (1992) reported that, estimate of D will be zero when the genes are equally dispersed among the parental lines, on the other hand the estimate will be greater than ∑d2 (d is the phenotypic differences between the two homozygotes) if the genes are associated. Thus, the measure of average degree of dominance, ½ (H1/D) , may either be inflated or reduced by the combined effect of correlation of genes on estimates of H1 and D and hence partial dominance may be converted into over dominance. ½ The (H1/D) ratio obtained in the material used, suggested that, the dominance component of genotypic ratio was more important, although both additive and non-additive components govern the expression of the character. The ratio of heritability was low indicating that large part of the variability for this trait was nonadditive in nature. However, in F1(2004), the additive variance was more important compared to the non-additive part and a moderate heritability estimate was observed (Table 5). Hence, mass selection is not likely to lead to much genetic improvement ( Gomathinayagam and Dabholkar,1992). El-Shazaly et al.(1995) concluded that, care should be taken when planning breeding programme for selection of thin seed coat, since the genetic behavior of the trait is related to the use of specific genetic material. Three to four genes were observed to control seed coat weight as indicated by the ratio ½ ½ [(4DH1)) +F]/[(4DH1) –F] . Among them one gene was dominant (h2/H2 approx. = 1). 5.1.2.2 (wr,vr) Graph In F1(2003) graph, (Fig 4), the regression line had a slope of 0.69±0.41. This value was not significantly different from zero or unity, indicating that the additive dominant model was satisfied in this trial. However, in F1(2004) and F2 (Figures 5 and 6) the regression lines slopes were 1.65±0.45 and 1.69±0.31, respectively. These values were significantly different from zero and unity indicating the presence of gene interaction in the material used (Hayman,1954 and Whitehouse et al (1958). The interception points were below the origin in the three graphs, indicating over dominance of the character in the experimental material. Hence, the results showed by the graphs (Figures 5 and 6) were different from that expressed by the estimates of the genetic ½ parameter ratios,(H1/D) , in that the mean degree of dominance in F1(2004) and F2 was less than one. In the diallel analysis, it is assumed that genes are independently distributed among parents, failure of this assumption may result either due to linkage of genes in the population from which parents were selected or due to the effect of sample size when choosing the parents (Baker,1978). Also Coughtrey and Mather (1970) postulated that, the (wr,vr ) graph disturbed due to non-independent distribution of genes. In all the graphs (Figures 4, 5 and 6), the array points scattered around the regression lines indicating their variability. The array points of 1(BB7) and 2(C.28) appeared to posses the least dominant alleles. These two parents showed either negative or the smallest Fr values (Appendix 5). On the other hand, array points 4(C.36/1) and 8(C.36) appeared to posses the most dominant alleles; these two parents posses either large or the largest positive Fr values. The array points 3(C.86/1), 5(ZBF/1/1), 6(Bulk/1/1/2) and 7(C.22) had changed their orders and they were middle in position. The material used in this experiment (which are from F6 and F7 generations) were chosen in 2001/2002 season according to their genetic variability in seed hardness and seed coat weight. Also, the evaluation in previous generations was concentrated mainly on their yield performance. Eberhart and Gardner (1966) expressed the view that, diallel parents are should be usually selected from material of interest to the breeder, and thus cannot be regarded as a random sample. Estimation of genetic variance components do not provide any useful information. 5.2 Combining Ability In this investigation, seed hardness showed significant specific combining ability (sca) in both seasons and general combining ability (gca) in the first evaluation season only. This difference may be due to the difference in value of error variance (Table 7). The contribution of gca and sca for both seed coat weight and 100-seed weight were significant indicating that both additive and nonadditive gene actions determine the expression of these traits. A similar result was obtained by Waly and Abdel Aal (1987). The non-additive genetic variance was more important than the additive genetic variance for number of pods/plant and number of seeds/plant indicating the dominant effect on both traits. Moreover, the gca/sca ratio confirm the importance of the non-additive effect (gca/sca ratio <1 for all traits). For seed yield/plot both components of genetic variance were nonsignificant. This reflects the importance of the effect of environment on yield. The results regarding yield and its components were in accordance with those reported by El-Hosary (1984,1985) and El-Hosary et al. (1997). Furthermore, El-Hosary et al. (1997) concluded that selection for these characters would be effective in early generations. Among the parents 2(C.28), 3(C.86/1) and 4(C.36/1) gave the desirable gca effect for all traits, except, in the parent the parent 4(C36/1) showed positive significant effect for seed coat weight in the second season (Table 8). Thus, they were good general combiners for these traits. For sca effect, in seed hardness, all crosses which exhibited the positive significant effect (1×3, 1×7, 2×5, 2×6, 3 ×5, 5×6 and 6×8) had one or both of their parents exhibiting positive gca effect, except the cross I×3, in which both parents showed negative effect. On the other hand, the crosses 1×6, 2×3, 2×7, 3×7, 5×7 and 5×8, which showed negative significant sca effect had one or both of their parents exhibiting positive gca effect except the cross 2×3 in which both parents showed significant effect for low percentages of hard seed. This either reflects the prevalence of the dominant genes in all parents used (Figures 1,2 and 3), due to the ambidirection of genes (b2 and H2 were significant, Tables 4 and 5) or the effect of the environment on this character was large. El-Hosary (1985) concluded that gca effects of the parental lines were generally unrelated to sca values of their corresponding crosses. Moreover, Bond (1967) and El-Hosary (1984, 1985) reported that parents with significant gca effects did not necessarily produce hybrids with high sca effects and vice versa. For seed coat weight, the crosses 1×3 and 4×7 showed significant negative sca effect. In these two crosses, the parents 1 ,3 and 4 showed desirable gca effect. On the other hand, the crosses 1×6 and 2×6 exhibited positive significant sca effect, in them the parents 1 and 2 gave high negative gca effect. Regarding the yield and its components, the crosses 4×8 and 5×6 showed significant positive sca effect but they exhibited significant effect towards the high percentages of hard seed. The crosses 2×3 and 5×8 had desirable sca effect for all characters. Both of the parents of the cross 2×3 were good combiners. However, the parents of cross 5×8 were undesirable combiners. If the cross exhibited high sca values and both of its parents also are good general combiners, they could be exploited for breeding varieties as well (El-Hosary, 1985). 5.3 Correlations The results obtained in the present investigation indicated that yield/plot exhibited significant association with number of pods/plant, number of seeds/plant and 100-seed weight, and the highest association was that with number of pods/plant, which was generally in the same line with what was reported in the literature. The correlations of 100-seed weight with number of pods/plant and number of seeds/plant were negative. This result of negative associations was confirmed by Kambal (1969); Yassin (1973); Magyarosi and Sjodin (1976); Mahmoud et al. (1978) and Neal and Mc Vetty (1984). The negative association of number of pods/plant with 100-seed weight is attributed to the competition for assimilates (Adam, 1967). The associations of seed hardness with yield and its components were not significant. This was in accordance with what was reported by Salih (1976) and Ramsay (1997). However, the seed coat weight exhibited significant correlation with yield in the second season only, which was in contrast with that reported in the literature. This contradicting result may be due to the large variability exhibited by the line used in the second season. 5.4 Effect of Pods Position on Seed Hardness and Seed Coat Weight a. Seed Hardness: The result presented on the effect of pod position on the plant for hard seed (Table 12 and Appendix 11) showed that lower pods gave high percentages of hard seeds than upper pods. In the first experimental season parents were screened as with high hard seeds percentages or with low hard seeds percentages. The same trend still was observed in the two groups. All the parents with the high hard seeded group gave high percentages in the lower pods and lower percentages in the upper ones and the difference between the two positions was large. However, the low hard seeded group showed inconstant trend (Appendix 11). Zenari (1929); Quinlivan (1965) and El- Bagoury (1975), reported that, seed dormancy depend on the extend of seed maturity i.e. increased markedly with delayed maturity. Thus, they assumed that long exposure to temperature during the growing period may affect the permeability of the seed coat. b Seed Coat Weight The pod location on the stalk had no effect on seed coat weight. This showed that, the genetic factors play important role in the expression of this trait. Rowland and Fowler (1977) found that the position of the seed on the plant had no effect on the seed coat thickness. 5.5 Seed coat structure Seed hardness in faba bean is attributed to many causes, among them the physical and morphological characteristics of seeds. The palisade layer attracts much attention and its structure is connected with the high degree of impermeability. A magnified view of the palisade cells (Fig 7 b) showed their fibrous nature. They are highly compact and devoted of intercellular spaces. The hard seeds showed small length and width compared to the soft seeds (Table 14). The hourglass cells showed a variety of sizes and shapes. The hard seeds measured small range compared to the soft seeds. The seed coat thickness get wider near to the micropyle region in the four groups. The hard seeds had small thickness compared to the soft seeds in the two lines (Table13). The micropyle opening is the only gate for water entery to the seed. In this region, the trachid cells are found (Fig 8 d ). Water and nutrients enter the maturing seeds through these trachids. Thus its width and length play an important role in water absorption. Esua (1979) reported that, this region acts like a hygroscopic valve. The two lines exhibited a clear difference in measurements (Table 16). The micropyle and trachids in the hard seeds got approximately near half that of the soft seeds in the two lines. Also line 8(C.36 ) which belongs to the hard seeded group, got small measurement compared to line 2(C.28) Youssef and Bushuk (1984) found that, the seeds of the hard - to cook Egyptian samples had smaller micropyle opening, shorter hourglass cells and thicker cell layers than those of the soft - to cook types. Similar observation was published for dry field bean (P. vulgaris L) by Powrie et.al. (1960). REFERENCES Abdalla,M.M.F. (1976). Natural variability and selection in some local and exotic populations of field beans (Vicia faba L.). Z. Pflanzenzenzuchty 76: 336-343. Abu El-Zahab, A. A.; A. A. Ashore nnd K. H. El-Hadedy (1980).Comparative analysis of growth, development and yield of five field bean cultivars (Vicia faba L.). Z. Acherund pflanzenbu 149: 1 – 13. Adam, M. W. (1967). Basis of yield components compensation in crop plants with special reference to field bean (Phaseoulus vulgaris L.). Crop Sci. 7: 505-510. Ageeb, O.A. A.(1980). Effect of harvest on grain yield and percentage of hard seed of faba bean (Vicia faba L.). FABIS 2: 38-39. Altikin, Yvorme (1939). The problem of hard seeds in Subterranean clover. Proc. Ray. Sac.Vict. 51: 187-193. Ayoub, A. T, (1971). Ful Masri (Broad bean) fertilizer experiment. Auual Report (1970/197)1, Hudeiba Res. Station. —————— (1973). Ful Masri watering experiment. Annual Report (1972/73), Hudeiba Res. Station. Baciu-Miclaus, D. (1970). Contribution to the study of the hard seed and seed coat structure properties of soybean. Proc. Int. Seed Test. Ass. 35: 366–617. Baker, R. J. (1978). Issues in diallel analysis. Crop Sci. 18: 533–536. Bakheit, R. B. and E. E. Mahdy (1988). Variation, correlation and path coefficient analysis for some characters in collections of faba bean (Vicia faba L.). FABIS 20: 9–13. Bassett, M, (1997). A new allel (vwf) at the V locus for flower and seed colour in common bean. J. of American Society for Hort. Sci. 122(4): 519–521. —————— (1998). Athird recessive allel, stomic, for seed coat pattern at the stp locus in common bean. J. of American Society for Hort. Sci. 123 (4): 404–406. Bell, J. M. (1975). Utilization of protein supplements in animal feeds. pp.633-668 in (J. T. Harapiak, ed.) Oil seed pulse crops in Wesyern Canada – Asymposium. Western Cooperative Fertilizers Ltd. Bennett, H. W. (1959). The effectiveness of selection for the hard seeded character in crimson clover. Agron. J. 51: 15–16. Bond, D. A. (1967). Combining ability of winter bean (Vicia faba L.) inbreds. J. Agric. Sci. Camb. 68: 179–185. Cerning, J. A.; A. Saposnik and A. Guilabot (1975). Carbohydrate composition of horse bean (Vicia faba L.) of different origins. Cereal Chemistry 52: 125–138. Coughtrey, A. and K. Mather (1970). Interaction and gene association and dispersion in diallel crosses where gene frequances are unequal. Heredity 25: 79–88. Crumpacker, D. W. and R. W. Allard (1962). A diallel cross analysis of heading date in wheat. Hilgardia 32(6): 275– 319. Dhirendra, Kare and C. B. Singh (1984). Inheritance of seed dormancy in Vicia faba L. FABIS 8: 4-5. Donnelly, E. D. (1971) Breeding hard seeded vetch using interspecific hybridization. Crop Sci. 11: 721–724. ————— ; J. E. Watson and John, A. Mc Guire (1972). Inheritance of hard seed in vicia. Heredity 4: 361–365. Eberhert, S. A. and C. O. Gardner (1966). A general model for genetic effects. Biometrics 22: 864–881. El-Bagoury, O. H. (1975). Effect of different fertilizers on the germination and hard seed percentage of broad bean seeds (Vicia faba L.). Seed Sci. Technol. 3: 569–574. El–Hady, M. M.; M. Omer; S. Nasri; K. Ali and N. Essa (1998). Gene action on seed yield and some yield components in F1 and F2 crosses among five faba bean (Vicia faba L.) genotypes. Bulletin of Faculty of Agriculture, University of Cairo 49(3): 369–388. El–Hosary, A. A. (1984). Heterosis and combining ability in diallel crosses among seven varieties of faba bean. Egypt. J. Agron. 9 (1–2): 17–28. ———— (1985). Heterosis and combining ability in F1 and F2 diallel crosses of faba bean (Vicia faba L.). Egypt. J. Agron. 10 (1–2): 13–24. ————— : S. Shoker; A. A. El–Halim; S. M. Nasr and A. M. El-Aziz (1997). Heterosis and combining ability in faba bean (Vicia faba L.) Egypt. J. Agric. Res. 75(3): 811–826. El–Mubarak, A. A.; A. F. Salih; A. N. El–Hud. and A. M. Gurashi (1988). Effect of time of harvest on physical and chemical composition, cookability and yield of faba bean. FABIS 20: 33– 36. Esau, K. (1977). Anatomy of seed plants. 2nd Ed. John Wiley and Sons, New York. El-Shazly, M. S. ; M. M. El–Ashry ; El-Sayed M. Gaafar and Magdy M. M. Mohamed (1995). Inheritance of some seed quality characters in faba bean. FABIS 36/37: 8–13. Falconer, D. S. (1980). Introduction to Quantitative Genetics. 2nd Ed. Longman, London. FAO (2002). Production Year Book 56: 111. FAO, Rome. Forbs, I. and D. H. Well (1968). Hard and soft seededness in blue lupine (Lupinnus angustifolius L.), inheritance and phenotype classification. Crop Sci. 8: 195-197. Gomathinayagam, P. and A. R. Dabholkar . (1992). Elements of Biometrical Genetics. Concept Publishing Company, New Delhi 110059. Griffing, B. (1959). Concept of general and specific combining ability in relation to diallel crossing system. Aust. J. Biol. Sci. 9 : 461-493. Habetinek, J. ; M. Ruzickoua and J. Soucek (1983). Variability of characteristic in and correlation between various cultivars of horse beans (V. Faba L.). Faba bean Abstracts .3(1): 2–2. Harrington, G. T. (1916). Agricultural values of impermeable seeds. J. Agric. Res. 6: 761–795. Hayman, B. I. (1954). I : The theory and analysis of diallel crosses. Genetics 39: 789–809. ——————— (1958). II : The theory and analysis of diallel crosses. Genetics 43: 63–85. Hussain, M. G. ; N. Hill and N. Gallage (1988). The response of field bean (Vicia faba L.) to irrigation and sowing date. 2: Growth and development in relation to yield. J. Agric. Res. Camb. 111: 233–254. Ikram, A. F. (1993). Genetic variability, correlations and genotype×environment interaction in faba bean (Vicia faba L.). M. Sc. U. of Kh. Jha, B. N. and R. P. Singha (1989). Hard seededness in Vicia faba L.. FABIS 24: 37–38. Jinks, J. L. (1954). The analysis of continuous variation in diallel crosses of Nicotiana rustica varieties. Genetics 39: 767–788. Johnson, H. W. ; H. F. Robinson and R. E. Comstok (1955). Genotypic and phenotypic correlations in soy bean and their implications in selection. Agron. J. 47: 477–482. Johnes, R. M. (1965). Analysis of variance of the half diallel table. Heredity 20: 117–121. Kambal, A. K. (1962). Astudy on combining ability in grain sorghum. Ph. D., U. of Kalifornia. —————— (1969). Components of yield in field bean (Vicia faba L.). J. Agric. Sci. Camb. 72: 359– 363. Kaul, D. K. and K. L. Vaid (1996). Combining ability in faba bean. FABIS 38/39: 12–14. Klasinsko, K. (2000). Seed sowing value and seed coat structure of bean (Phaseolus vulgaris L.). Biuletin Institute. Hodowli. 2– Aklimatyzoeji Roslin 214: 245–252. Lebedeff, G. A. (1947). Studies on the inheritance of hard seed in beans. J. Agric. Res. 74: 205–215. Lute, A. M. (1928). Impermeable seed of alfalfa. Colorado Agri. Exp. Sta. Bull. 326. Magyarosi, T. and J. Sjodin (1976). Investigation of yield and yield components in field bean (Vicia faba L.) varieties with different ripening time. Z. Pflanzenzutechty 77: 133–144. Mahmoud, S. A. and El-Ayoubi (1986). Seed yield and seed size in diallel crosses between some cultivars of broad bean (Vicia faba L.). Bulletin of Faculty of Agriculture, Cairo Univ. 37(2): 603–612. ——————; Correlation and Y. El–Hyatmey and F. M. El Rayse (1978). and path coefficient analysis of yield components chemical composition in broad bean (Vicia faba L.). Agricultural Research Review (Egypt). 56(8): 117–125. Marcellos, H. (1987). Variation in testa fraction with seed weight in faba bean. FABIS 18: 33–34. Mather, K. and J. L. Jinks (1949). Biometrical Genetics. Chapman and Hall Ltd. London. EC4. Mayer, A. N. and A. P. Mayber (1982). Germination of Seeds. 3rd. Ed. Pergamon Press. New York, USA. pp. 53. Mc Ewen, T. J.; B. L. Dranzek and W. Sushuk (1974). Scanning electron microscope study of faba bean seed. Cereal Chem. 51: 750–757. Mutwakil, A. M. (1965). Ful Masri variety trial at Hudiba and Shendi. Annual Report (1964/65), Hudeiba Res. Station. Naidu, M. R.; C. Ashok and V. P. Singh (1985). Analysis of yield components in broad beans. Indian J. Agric. Sci. 55(4): 236–239 Neal, J. R. and P. B. Mc Vetty (1984). Yield structure of faba bean (Vicia faba L.) grown in Manitoba. Fiela Crop Res. 8: 349– 360. Ojomo, O. A. (1972). Inheritance of seed coat thickness in cow peas. J. Heredity 63: 147–149. Picard, J. and P. Berthelem (1980). Abrief comment on yield stability and thousand grain weight in Vicia faba L. . FABIS 2: 20–20. Povilaits, B. (1970). Diallel Analysis of crosses between Flue cured and barley Tobacco cultivars. Can. J. Genet. Cytol. 12: 484– 489. Powerie, W. D.; M. W. A. Adams and I. J. Pflug (1960). Chemical studies on the navy bean seed. Agric. J. 52: 163–169. Quinlivan, B. J. (1965). The influence of the growing season and the following dry season on the hard seededness of subeeanean clover in different environments. Aust. J. Agric. Res. 16: 277–291. —————— (1968). The softening of hard seeds of sand-plain lupine (Lupinus varuis L.). Aust. J. Agric. Res. 19: 507–515. Rajendra, B. R.; K. A. Mujeeb and L. S. Bates (1979). Genetic analysis of seed coat types in interspecific vigna hybrids via SEM. J. Heredity 70: 242–249. Ramsay, G. (1997). Inheritance and linkage of a gene for testa – imposed seed dormancy in faba bean (Vicia faba L.). Plant Breeding 116(3): 287–289. Rowland, G. G. (1970). Seed coat thickness and seed crude fibre in faba bean (Vicia faba L.). Can. J. Pl. Sci. 57: 951–953. ————— and D. B. Fowler (1977). Factors affecting selection for seed coat thickness in faba bean (Vicia faba L.). Crop Sci. 17: 88–90. Saeed, E, M.; S. O. Fregoun; M. E. Omer and S. B. Hanounik (1987). Effect of wilt and root rot disease complex on some quality parameters of faba bean (Ful Masi) 23–26. seeds. FABIS 19: Salih, F. A. (1976). Broad bean standard variety trial. Annual Report (1975/76), Hudeiba Res. Station. ————— (1978 a). Broad bean water closure experiment. Anuual Report (1977/78)., Hudeiba Res. Station. ————— (1978 b). Field beans time of harvest. Annual Report (1977/78), Hudeiba Res. Station. ————— (1979 a). Effect of sowing date, seed size and seed rate on seed yield of broad bean. Annual Report (1978/79), Hudeiba Res. Station. ————— (1979 b ). Effect of different nitrogen and phosphorous levels and different methods of application on broad bean yield and its components. Annual Report (1978/79), Hudeiba Res. Station. ————— (1979). Hard seed problems with Vicia faba in the Sudan. FABIS 1: 33. ––––––––– (1981). Breeding work on Vicia faba in the Sudan. FABIS. 3: 21–23. ————— (1982a). Influence of seed size and sowing date on yield and yield components of faba bean. FABIS 5: 13–14. ————— (1982 b). Hard seeds in faba bean. In: Faba Bean Improvement. (Ed. G. Hawtin and G. Webb). pp. 363–373, Martinus Nijhaff, for the ICARDA/IFAD Nile Valley Project. ————— (1983a). Breeding work on Vicia faba in the Sudan. In: More Food for Better Technology. (Ed. Holmes, J. C. and W. M. Tahir). pp. 768–777. FAO, Rome, GCP/INT /387/SW. —————— (1983b). Broad bean (Vicia faba L.) grain yield and percentage of hard seed as affected by time of harvest. J. Agronomy and Crop Sci. 152: 394–398. —————— (1987). Effect of nitrogen application and plant population per hill on faba bean (Vicia faba L.) yield. FABIS 17: 27–30. —————— and A. E. Ali (1986). Effect application and time of harvest on seed yield of phosphorous and quality of faba bean. FABIS 15: 32–35. —————— and N. E. A. Khairi (1990). Variation in testa fraction with some other seed quality attributes of faba bean grown in the new production areas in the Sudan. FABIS 27: 30–31. Salih, S. H. (1995). Faba bean Research in the Sudan, an over view . A paper presented in Rehabilitation of Faba Bean Seminar in Rabat, Moroco,24–26 May. ————— and F. A. Salih (1985). Faba bean breeding, Back-up research. ICARDA/IFAD NVP six Year Report. ————— and ————— (1996). Faba bean Improvement .In: Production and improvement of Cool Season Food Legumes in the Sudan (Ed. Salih, S. H.; A. O. Ageeb; C. M. Saxena and B. M. Salih). pp. 19–32, Proceeding of the National Research Review workshop, 27–30 August, 1995, Wad Medani, Sudan. Sass, John, E. (1958). Botanical micro-technique. The Lowa State, University Press. Shalaby, I. A. and Y. S. Katta (1976). Path coefficient analysis of seed yield and some agronomic characters in field beans (Vicia faba L,). J. Agric. Res. (Egypt) 2 (2): 70–79. Shaudary, K. A. and G. H. Buth (1970). Seed coat structure and anatomy of Indian pulses. J. Linn. Soc. Soppl. 1(63): 169–179. Shehata, A. M. E.; H. M. Zeinah and M.M. Youssef (19870). Effect of seed coat cotyledon on the structure of cooked faba bean (Medammis). FABIS 19: 20–22. Sindhu, J. S.; O. P. Singh and R. P. Singh (1985). Selection indices in faba bean (Vicia faba L.). FABIS 12: 7–7. Singh, R. . and B. D. Shaudary (1977). Biometrical Methods in Quantitative Genetic analysis. Kalyani Puplishers. New Delhi, Ludhlana. Spraque, G. F. and L. A. Tatum (1942). General vs. specifc combining ability in single crosses of corn. J. Am. Soc. Agron. 34: 923–932. Waly, E. A. and S. A. Abdel Aal (1987). Inheritance of seed coat weight in faba bean (Vcia faba L.). FABIS 18: 4–5. Whitehouse, R. N .H.; J. B. Thompson and M.A.M. Dovalleribeiro (1958). Studies on the breeding of self- pollinating cereals. 2: The use of a diallel cross analysis in yield production. Euhpytica 7: 147–169. Yassin, T. E. (1973). Genotypic and phenotypic variances and correlations in field beans (Vicia faba L.). J. Agric. Sci. Cam. 80: 445–448. Youssef M. And M. Bushuk (1984). Microstructure of the seed coat of faba bean (Vicia faba L.) seeds of different cookability. Cereal Chem. 61(4): 381–383. ————— ; —————; E. D. Murray and A.M. El–Iabeshchata (1982). Relationship between cookability and some chemical and physical properties of faba bean (Vicia faba L.). J. of Food Sci. 47: 1695. Zenari, S. (1929). The character hard seed in relation to heredity and environment. Ann. Bot. 18: 174–215.






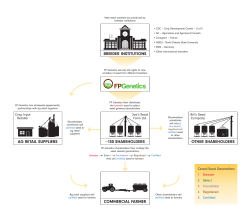



© Copyright 2026