
Hyperimmune globulins in prevention and treatment of respiratory syncytial virus infections.
Hyperimmune globulins in prevention and treatment of respiratory syncytial virus infections. V G Hemming, G A Prince, J R Groothuis and G R Siber Clin. Microbiol. Rev. 1995, 8(1):22. These include: CONTENT ALERTS Receive: RSS Feeds, eTOCs, free email alerts (when new articles cite this article), more» Information about commercial reprint orders: http://journals.asm.org/site/misc/reprints.xhtml To subscribe to to another ASM Journal go to: http://journals.asm.org/site/subscriptions/ Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest Updated information and services can be found at: http://cmr.asm.org/content/8/1/22 CLINICAL MICROBIOLOGY REVIEWS, Jan. 1995, p. 22–33 0893-8512/95/$04.0010 Copyright q 1995, American Society for Microbiology Vol. 8, No. 1 Hyperimmune Globulins in Prevention and Treatment of Respiratory Syncytial Virus Infections VAL G. HEMMING,1* GREGORY A. PRINCE,2 JESSIE R. GROOTHUIS,3 AND GEORGE R. SIBER4 Department of Pediatrics, Uniformed Services University of the Health Sciences, Bethesda, Maryland 208141; Virion Systems, Inc., Rockville, Maryland2; Department of Pediatrics, University of Colorado School of Medicine, and the Children’s Hospital, Denver, Colorado3; and Massachusetts Public Health Biologic Laboratories and Division of Infectious Diseases and Dana-Farber Cancer Institute, Boston, Massachusetts4 This review will discuss the initial recovery, the characterization, the epidemiology, and the impact on human populations of RSV. It will examine evidence from animal and human studies indicating protective and therapeutic roles for the parenteral administration of human polyclonal RSV-specific immunoglobulin G (IgG) antibodies (RSVIG), and perhaps RSV-specific monoclonal antibodies, for the prevention, amelioration, or treatment of RSV infections. Immunoprophylaxis provides a new clinical approach to the prevention and perhaps the treatment of RSV infections in certain high-risk infants and young children. Finally, studies indicating that neutralizing anti-RSV IgG antibodies introduced into the airway by inhalation may be an effective alternative to the parenteral injection of RSVIG for the treatment of RSV LRI in infants and children will be summarized. INTRODUCTION Respiratory syncytial virus (RSV) was first recovered from primates in 1955 (96) and humans in 1956 (21, 22). By 1961, it was evident that RSV was a frequent and sometimes serious respiratory pathogen for infants and young children. A formalin-inactivated vaccine was prepared and tested to determine whether vaccination might protect infants and young children from RSV lower respiratory tract infections (LRI) (28, 44, 79, 81). The vaccine stimulated a humoral antibody response. However, some of the youngest vaccine recipients later became infected with RSV, developed severe bronchiolitis, and required hospitalization. The investigators suggested that humoral antibody in the absence of mucosal immunity permitted the development of immune complex disease, which in turn provoked a more severe bronchiolitis. The acceptance of this hypothesis hampered scientific inquiry into a clearer understanding of the role of humoral immunity in the pathogenesis of infant RSV pulmonary infections. Subsequent evidence derived from human (17, 49, 72, 84, 143) and animal (69, 114–116, 119–120, 130) studies indicated that sufficient serum antibody was safe and, furthermore, protected the lower respiratory tract, airways and lung parenchyma, from RSV infection. RSV: AN HISTORICAL OVERVIEW Isolation and Characterization In October 1955, an apparent epizootic of sneezing, coughing, and mucopurulent nasal discharge occurred in a group of young chimpanzees housed at the Forest Glen facility of the Walter Reed Army Institute of Research in Washington, D.C. Investigators (96) cultured nasal secretions from symptomatic chimpanzees in human liver epithelial cells (Chang) and recovered a hitherto unrecognized virus which they called the chimpanzee coryza agent (CCA) and which they showed to be causally related to the monkeys’ upper respiratory tract infec- * Corresponding author. Mailing address: Department of Pediatrics, Uniformed Services University of the Health Sciences, 4301 Jones Bridge Road, Bethesda, MD 20814. Phone: (301) 295-3391. Fax: (301) 295-3898. 22 Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest INTRODUCTION .........................................................................................................................................................22 RSV: AN HISTORICAL OVERVIEW ........................................................................................................................22 Isolation and Characterization ...............................................................................................................................22 Epidemiology and Clinical Syndromes ..................................................................................................................23 Vaccine Development................................................................................................................................................23 Animal Models of RSV Infection............................................................................................................................24 Phenotypic Variation ................................................................................................................................................24 RSV AND HUMORAL IMMUNITY ..........................................................................................................................24 Evidence from Studies in Humans.........................................................................................................................24 Evidence from Studies in Animals .........................................................................................................................24 STUDIES OF STANDARD IVIG AND RSVIG FOR THE PREVENTION AND TREATMENT OF RSV INFECTION IN HUMANS..................................................................................................................................25 Standard IVIG...........................................................................................................................................................25 Development of RSVIG ............................................................................................................................................26 RSVIG Studies in Cotton Rats ...............................................................................................................................26 RSVIG Studies in Children .....................................................................................................................................26 TOPICAL OR INHALED HUMAN IG AND RSVIG STUDIES............................................................................28 IVIG Topical Studies in Cotton Rats and Owl Monkeys....................................................................................28 RSVIG Topical Studies in Cotton Rats .................................................................................................................29 RSV AND MONOCLONAL ANTIBODIES ..............................................................................................................30 CONCLUSIONS ...........................................................................................................................................................30 REFERENCES ..............................................................................................................................................................30 VOL. 8, 1995 HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS Kravetz et al. (82) and Johnson et al. (76) were able to infect the upper airways of adult volunteers with RSV despite the presence of serum RSV NT antibodies at the time of challenge. In contrast to the high seroprevalence found in adults, surveys of children showed RSV titers in only 35% of children under 4 years of age and 29% of those between the ages of 4 and 8 years. In retrospect, it is evident that the assays used in these pioneering studies were not optimally sensitive; however, further evidence of the high frequency of RSV infections in infants and children was provided. Epidemiology and Clinical Syndromes Other investigations (26, 57, 63, 64, 80, 85, 87, 94, 121) affirmed the initial observations regarding the epidemiology of RSV and its importance in human respiratory infections. The virus caused annual winter RSV epidemics. The resulting clinical syndromes of infected infants and young children included mild upper respiratory tract infections but also bronchitis, bronchiolitis, bronchopneumonia, and laryngotracheitis. These LRI syndromes could be particularly severe in infants under 6 months of age. Immunity following primary RSV infection was often insufficient in magnitude or duration to protect from reinfection in subsequent years. Reinfections were usually less severe and were more likely limited to the upper respiratory tract, especially in children older than 2 years. Most young infants with RSV infections had preexisting maternal antibody (10, 15, 24, 46, 63, 64, 76, 87, 90, 121, 132). Most humans experience RSV infection by the second year of life. Annual seasonal epidemics occur in most communities. When the virus is highly prevalent, most infants, children, and adults are placed at risk for infection or reinfection. One of the paradoxes of RSV biology is the failure of primary and repeat infections to provoke solid or durable immunity, especially of the upper respiratory tract (9, 50, 61, 72, 97). In addition to occasional serious infections in healthy infants and children (83, 85), other groups at risk for complicated RSV infections include hospitalized premature infants (15, 58, 59), premature infants discharged home during the RSV season (35, 53), hospitalized children (58, 129), infants and children with chronic cardiac (24, 85, 86, 93, 105) or pulmonary (1, 53, 105) disorders, immune system-compromised children and adults (20, 37, 38, 40, 60, 65, 73, 127, 133), and the elderly (2). Vaccine Development Clearly, RSV was a common and often serious winter lower respiratory tract pathogen for the very young. The elusive search for an effective RSV vaccine was initiated. In the early 1960s, a formalin-inactivated vaccine was prepared and tested in infants (28, 44, 79, 81). The Bernett strain of RSV, previously isolated in human embryonic kidney cultures, was propagated in human embryonic kidney cells and multiply passaged in vervet monkey kidney cells. Harvests of RSV-infected cells were clarified by low-speed centrifugation, inactivated with 1:4,000 formalin, concentrated 25-fold by ultracentrifugation, and further concentrated 4-fold by alum precipitation. This preparation is known as lot 100. Infants at four sites (two sites in Washington, D.C., and military-dependent children at Lowry Air Force Base, Colo., and Fort Ord, Calif.) each received three intramuscular doses of vaccine at approximately 1-month intervals. In one Washington, D.C., study site, nonvaccinated children were observed as controls. In the other Washington, D.C., site, control infants received a univalent, formalininactivated parainfluenza virus type 1 vaccine. Control infants in Colorado and California were immunized with a formalininactivated, aqueous trivalent parainfluenza virus vaccine con- Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest tions. Seed virus prepared from the original CCA isolation induced infection and similar respiratory symptoms in other young chimpanzees. CCA failed to induce detectable infections in guinea pigs, mice, rats, or chicken embryos. The unintentional spread of CCA was observed between infected and uninfected chimpanzees housed in a common facility. A laboratory worker also appeared to become infected with CCA after contact with infected chimpanzees. Random human serum samples were surveyed by complement fixation (CF) and virus neutralization for CCA immunity. The assays suggested that some humans (1 of 12 infants and children 6 months to 2 years, 0 of 12 children 3 to 6 years, 0 of 9 children 7 to 9 years, 2 of 12 children 10 to 14 years, 3 of 13 children 15 to 18 years, and 8 of 40 persons older than 18 years) had been infected in the past with CCA or a related virus (96). Shortly after the recovery of CCA, Chanock and colleagues (21, 22) isolated a CCA-like virus from an infant with bronchopneumonia (Long agent) and another from an infant with bronchiolitis (Snyder). Serologically and phenotypically, as characterized by the viruses’ cytopathic effect on cells in culture and by their neutralization by anti-CCA animal sera, the Long and Snyder agents appeared to be the same as CCA. Particularly striking was their ability to induce syncytia and multinucleated giant cells in Chang liver epithelial cells and KB cell cultures. It was suggested that respiratory syncytial virus might be a more suitable name than CCA. CF and RSV neutralization tests (NT) performed on convalescent-phase sera from infants with LRI were compared with those done on sera from uninfected age-matched controls. Test results indicated that many infants and children developed humoral immunity to RSV during the 5 months of the study. By 3 to 4 years of age, the majority of children studied (77 to 80%) possessed serum NT antibody (21). These observations were confirmed by Beem and colleagues (11), who cultured the upper respiratory tracts and tested sera from inpatient and outpatient infants and children with respiratory infections during the winter of 1959 to 1960 in Chicago, Ill. Two CCA-like agents which caused extensive syncytium formation and the development of multinucleated giant cells in HEp-2 cells were isolated. Over a 5-month period, similar agents were recovered from 41 other patients mostly with LRI. Thirty-one isolations were from children under 2 years of age, most of whom were less than 6 months old. Convalescentphase sera from the infected infants exhibited poor CF and/or NT responses to infection, whereas most older children showed significant antibody responses. Though a variety of clinical disease patterns were observed, the most common were bronchiolitis and bronchopneumonia. Prospective efforts (23, 108) to recover RSV from young patients in Washington, D.C., began in 1959. The first RSV isolation occurred in March 1960 during an epidemic of bronchiolitis. Subsequently, RSV was recovered from 57% of young infants with bronchiolitis or pneumonia during a 5-month period. RSV was also recovered from older children with bronchopneumonia and bronchiolitis. The frequent recovery suggested that RSV was an important human respiratory pathogen of early life. Serological data confirmed a high prevalence of RSV. Its epidemiology was different from those of influenza viruses A and B and parainfluenza virus types 1 and 3. Serious illness with RSV was most likely to result with the first infection, especially if it occurred in the early months of life. Reinfection appeared to be common but usually induced less severe illness in normal children. This appeared to explain why older children with nosocomial RSV infections, who usually were experiencing recurrence, became less ill than infants with primary infection. 23 24 HEMMING ET AL. Animal Models of RSV Infection Models of RSV infection in animals have contributed important information to the understanding of RSV pathogenesis and immunity. Cotton rats (Sigmodon hispidus) and small primates such as the owl monkey (Aotus trivirgatus) have been particularly valuable. Data from animal experiments have provided important information for use in vaccine development (13, 51, 78, 100, 102, 103, 110, 118–120, 122, 123, 130, 136, 148, 152). Contrary to expectations posited by the hypothesis suggesting that RSV antibody is harmful, passively immunized, RSVinfected animals such as cotton rats failed to demonstrate that passively acquired antibody led to enhanced disease. Indeed, the converse was observed. Passive antibody in sufficient titer prevented RSV LRI and also could be used to treat RSVinfected animals and reduce RSV titers in infected lung (52, 56, 111, 119, 130). Phenotypic Variation Phenotypic and serological variability was noted among RSV strains shortly after the original isolation and characterization (29, 30, 74, 151). With the biochemical characterization of the structure of RSV (31, 39) and the advent of monoclonal antibody technology (7, 8, 32), it became possible to characterize RSV strains (7, 41, 45, 47, 48, 77, 99). Grouping was based on substantial genetic heterogeneity (19, 33, 77, 131), especially involving the G glycoprotein, a protein putatively responsible for RSV attachment to cells. Antigenic variability of RSV strains and its possible role in explaining the phenomenon of reinfection (95, 96, 139, 144, 147), apparent differences in disease severity (62, 88, 89, 98), and RSV epidemiology (4, 126, 128, 137) are under investigation. RSV AND HUMORAL IMMUNITY Evidence from Studies in Humans Chanock et al. (23) and Beem (9) observed that young infants with RSV infections had NT titers to RSV at the time of infection. Some had ‘‘relatively high’’ titers and yet experienced bronchiolitis or bronchopneumonia. Beem reported that five of seven infants with documented repeat RSV infection had serum NT activity at the time of reinfection. RSV antibodies appeared to be poorly protective. It was also uncertain whether reinfections resulted from antigenic differences between infecting strains. In 1976, Lamprecht and coworkers (84) observed that, although maternally derived RSV NT antibodies did not prevent RSV infection in infants, the severity of RSV pneumonia correlated inversely with the level of NT antibody. This was not true with regard to the severity of bronchiolitis. In 1977, Bruhn and Yeager (17) reported no correlation among CF titers in cord blood, acute sera, or convalescent sera in 41 RSV-infected infants. An infant’s ability to respond with a fourfold rise in antibody titer after RSV infection appeared related to the amount of passively transferred maternal antibody. Infants under 2 months of age were relatively spared from infection, indirectly suggesting that antibody might be protective rather than harmful. Henderson and coworkers (72) reported a 10-year longitudinal study of respiratory infections in a small group of healthy children monitored from birth. During RSV epidemics, primary infection rates of these children exceeded 90%. The rate for second infections was reduced and that for third infections was reduced further. Infections induced NT responses but did not provide long-term protection in all children. Amelioration of illness severity resulted from successive RSV infections. Similar observations were made by Ogilvie et al. (106) and Ward et al. (143), who prospectively studied 100 newborns for evidence of RSV infection. Maternal antenatal sera were also examined for anti-RSV IgG by immunofluorescence and by radioimmunoprecipitation analysis. Twenty-nine of the infants developed RSV infection. The mean titer of maternal antibody in mothers whose infants remained uninfected was significantly higher than that of the mothers whose babies had proven RSV infection before 6 months of age. Glezen et al. corroborated these observations (49, 50). They observed that NT titers in cord sera of 68 infants with culture-proven RSV infection were significantly lower than those of 575 randomly selected cord samples of infants born during the same period. During prospective studies of primary RSV infections and reinfections in children observed from birth to 36 months of age, more than three-quarters of the infants and children experienced RSV infection in the first 2 years of life. Reinfection was common. The risk of reinfection correlated with the serum neutralizing antibody titer from the prior RSV infection and with the number of prior RSV infections. Evidence from Studies in Animals In 1975, Prince immunized pregnant female ferrets (111, 119) with live RSV prepared in HEp-2 cells. Infants born to RSV-inoculated mothers were challenged with RSV shortly after birth. Rather than experiencing enhanced disease, the neonatal ferrets were protected from RSV infection by transferred maternal immunity, and protection correlated inversely with the NT titers in their mothers’ sera (130). Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest taining types 1, 2, and 3. Serological studies of the vaccinees verified immune responses by increases in CF and NT antibody titers. When RSV appeared in the winter season following immunization, vaccinees were poorly protected from infection. RSV-exposed vaccinees became infected at the same rate as controls, and many experienced severe enough bronchiolitis to require hospitalization. Two deaths occurred in RSV-infected vaccinees. Formalin-inactivated RSV vaccine not only failed to protect children from RSV infection but appeared to induce an exaggerated, altered clinical response to naturally occurring infection (28, 44, 79, 81). To explain the unexpected outcome of the trial, it was proposed that vaccine-induced RSV serum antibody (and similarly, transplacentally acquired maternal antibody in nonvaccinated infants under 6 months of age), in the absence of local or mucosal RSV immunity, promoted an immunologic reaction that was responsible for severe pulmonary disease (25, 27). Similarities between the untoward outcome of the formalin-inactivated RSV vaccine trials and the complications observed in some recipients of a formalininactivated measles vaccine were noted. Both vaccines were prepared by the same manufacturer in a similar fashion, i.e., grown in simian kidney cells, formalin inactivated, and alum precipitated. An important difference in outcome between the two vaccine trials was noted. The measles-immunized children were initially protected from measles infection and then later became susceptible and, with infection, developed an altered disease state known as atypical measles (42, 43, 104). The fact that natural infection confers a measure of protection against severe repeat RSV infection justifies the continued efforts to develop vaccines (12, 14, 16, 18, 75, 91, 102, 107, 120, 138, 140–142, 145, 146, 149, 150). To date, the immune responses of infants and young children, immunized with investigational live or subunit vaccines, have provided neither sufficient nor durable protection. CLIN. MICROBIOL. REV. VOL. 8, 1995 HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS 25 TABLE 1. Effect of passive immunotherapy on amount of virus present in the nose and lungs of owl monkeys 5 and 7 days postinfectiona Virus titer (log10 PFU/ml) 5 days postinfection Treatment (n) Lung None (8) 3.42 6 0.22 IVIG (12) 3.02 6 0.18b 7 days postinfection No. (%) of monkeys without detectable virus Nose No. (%) of monkeys without detectable virus Lung No. (%) of monkeys without detectable virus Nose No. (%) of monkeys without detectable virus 0 (0) 0 (0) 3.37 6 0.31 3.67 6 0.22b 0 (0) 0 (0) 3.20 6 0.37 1.48 6 0.73c 0 (0) 9 (75) 3.32 6 0.40 2.10 6 0.33d 1 (13) 5 (42) a Titers are expressed as geometric means 6 standard errors. Significance was determined by comparison with untreated monkeys on the same day. Reprinted from reference 69 with permission of the publisher. b P . 0.05, not significant. c P , 0.001, significant. d P , 0.05, significant. immunized animals developed pulmonary inflammatory lesions within 24 h which peaked in severity 4 days after infection. The examination of microscopic sections of involved lungs revealed lesions that resembled pulmonary Arthus reactions (neutrophilic infiltration) at 24 h and lymphocytic peribronchiolitis at 96 h. Control animals similarly immunized with doses of heat-killed RSV demonstrated no pulmonary abnormalities. These findings suggest that the inflammatory response represents an artifact of formalin inactivation of RSV (117). These observations became extraordinarily important when we proposed to use exogenous human polyclonal antibodies for the prevention or treatment of RSV infections in humans. The success of IVIG prophylaxis and therapy in cotton rats prompted our testing of the safety and efficacy of IVIG for the treatment of RSV infections in primates. Adult owl monkeys were inoculated intratracheally with RSV (69). Bronchoalveolar lavage and nasal swabs were performed every other day from the 3rd to the 14th infection day in each monkey. Quantitative RSV cultures were performed on each swab and bronchoalveolar lavage specimen. On the fifth infection day, 12 animals were infused intravenously with a high-titered lot of IVIG. Eight other infected, but untreated monkeys served as controls (Table 1). IVIG treatment on the fifth day of infection (3,000 mg/kg of body weight given in three divided doses over 24 h) resulted in a mean (approximate) 30-fold reduction (P , 0.001) in virus from bronchoalveolar lavage fluid. There was complete clearance of RSV from bronchoalveolar lavage fluid in 9 of the 12 monkeys 48 h after treatment. RSV was also significantly reduced in the noses of the monkeys (69). STUDIES OF STANDARD IVIG AND RSVIG FOR THE PREVENTION AND TREATMENT OF RSV INFECTION IN HUMANS Standard IVIG The previously cited antibody data and the observations in cotton rats and owl monkeys prompted an examination of parenteral infusion of IVIG for the treatment or prophylaxis of RSV infections in young children. It appeared that standard commercial IVIG lots might be screened by RSV NT testing and high-titered lots could be selected for use in the immunologic mediation of RSV infections in children. Lots were screened and a high-titered lot of Sandoglobulin was identified. This screened lot was used (2,000 mg/kg given over 24 h) in a blinded, placebo-controlled trial to treat infants and young children hospitalized with RSV disease (71). The pilot study enrolled 35 hospitalized infants and children Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest Dreizin et al. (36) reported that adult cotton rats were permissive for RSV infection. Prince and coworkers have used the cotton rat extensively to study RSV pathogenesis and immunity (115, 116, 118). While ferrets are susceptible to pulmonary RSV infection up to the time of adolescence, cotton rats are susceptible throughout life. Histopathologic changes in cotton rats include rhinitis of moderate severity, mild bronchiolitis, and pneumonitis. Convalescent animals develop protective NT antibodies and cellular immunity that protects the upper airway and the lung from subsequent infections. Intramuscular immunization of adult nonimmune cotton rats with live RSV provides solid immunity for immunized mothers and their offspring. Immune mothers transfer immunity to their offspring mostly in colostrum and milk. Infants of immunized mothers respond poorly to subsequent RSV immunization as long as sufficient maternal immunity persists. Animals that are convalescent from RSV infections develop complete resistance to pulmonary reinfection which lasts at least 18 months, but nasal immunity is diminished by 8 months after infection. Passive immunization with convalescent-phase cotton rat serum given intraperitoneally to infant animals confers little nasal protection yet provides solid pulmonary protection from RSV infection. The level of serum NT antibodies required to confer pulmonary protection in the cotton rat (116) is similar to the NT antibody levels reported by Glezen et al. (49) to be protective for human infants under 2 months of age. Some lots of commercial standard human immunoglobulin prepared for intravenous administration (IVIG) contain substantial levels of NT antibodies to RSV (66). Several human IVIG lots were examined for prophylactic and therapeutic anti-RSV activity in inbred cotton rats (114). The prophylactic and therapeutic effects of four commercial lots of IVIG (Sandoglobulin; Sandoz, Inc., East Hanover, N.J., NT titers between 2,702 and 9,344) were compared with a high-titered human serum (NT titer, 2,800), convalescent-phase cotton rat serum (NT titer, 1,280), and normal nonimmune cotton rat serum (NT titer, ,20). In all cases the levels of RSV replication were inversely proportional to the NT titers achieved in the serum of the passively immunized cotton rats (114). It is noteworthy that animals treated therapeutically had a depressed primary antibody response to infection but were resistant to reinfection with RSV (114). The cotton rat model of RSV infection also provided a means to investigate potentiation of RSV infections by immunization with formalin-inactivated vaccines (117). Cotton rats were immunized with three doses of the original lot 100 vaccine or with similarly prepared formalinized vaccines. Convalescent animals were challenged intranasally with RSV. The 26 HEMMING ET AL. Development of RSVIG Investigators at the Massachusetts Public Health Biologic Laboratories and MedImmune, Inc. (Rockville, Md.) collaborated to develop RSVIG for use in the pending formal immunoprophylaxis efficacy trial (125). During the development of RSVIG, several techniques for screening individual plasma units for RSV-specific antibodies were examined. For the efficient and cost-effective production of RSVIG from pooled human plasma, it is necessary that the screening process use minimal amounts of plasma. The screening technique must also be amenable to automation. Intuitively, it was expected that enzyme-linked immunosorbent assay (ELISA) would be the simplest screening method and the easiest to automate. Several assays were tested: (i) an ELISA using whole-virus antigens from lysates of RSV-infected HEp-2 cells (125), (ii) direct ELISAs with purified RSV F and G glycoproteins as the solid phase (101, 140, 142), (iii) a competitive ELISA with monoclonal antibodies to the F glycoprotein (3, 6), (iv) live RSV plaque reduction assays with and without addition of exogenous complement (118), and (v) a live RSV microneutralization assay (5). Plasma pools were prepared from ‘‘high-titered’’ units identified by each of the methods. Each plasma pool was tested in a series of blinded, controlled, in vivo experiments of RSV prophylaxis in BALB/c mice. The subsequent analysis showed that the microneutralization assay best identified the plasma pools that protected the animals from RSV infection. Use of the assay to select individual plasma units for inclusion in a pool enhanced the in vivo NT activity of RSVIG by severalfold compared with standard IVIG. The enhanced activity was documented in the cotton rat (124) and in RSV immunoprophylactic studies in young children (55). RSVIG Studies in Cotton Rats Figure 1A and B depicts the relationship between cotton rat serum RSV NT activity, as determined by complement-enhanced plaque reduction, and the amounts of RSV recovered from lung or turbinate homogenates of infected animals. The serum neutralizing antibody (A/Long Potash strain) titer required to reduce RSV concentration in the lung by 99% was 390. To achieve a similar reduction in the nose required an NT titer of 3,500 (124). Figure 2 shows the serum RSV NT titers achieved (6 the standard error of the mean) in groups of cotton rats inoculated intraperitoneally 1 day before RSV challenge with 0.5 g of randomly selected lots of standard IVIG (n 5 24 animals) or RSVIG (n 5 47 animals) per kg of body weight or similarly inoculated with 5.0 g of either the standard IVIG (n 5 20 animals) or RSVIG (n 5 52 animals) per kg. Figures 3 and 4 compare the titers (log10 per gram of tissue) of RSV recovered from the lungs or turbinates of each group of cotton rats when the animals were euthanized 5 days after passive immunization and of unimmunized controls 4 days following RSV challenge. As was observed in previous cotton rat prophylaxis experiments, RSV NT antibodies protect the lower airway better than the nose and turbinates. Significant differences in the protective effect are noted between standard IVIG and RSVIG (P , 0.01) in both the nose and lung. The treatment outcomes in these studies correlate directly with the NT antibody titers in the treatment lot or with the dose of IVIG (or RSVIG) administered (124). RSVIG Studies in Children Groothuis and the RSVIG Study Group (54) demonstrated that infants and children with BPD or congenital heart disease and infants born prematurely in the months immediately preceding the RSV infection season tolerated the once-monthly infusion of IVIG in the range of 500 to 750 mg/kg. Consequently, a prospective three-arm, blinded, randomized, controlled trial of RSVIG was implemented. A placebo arm was not included in the trial because it was deemed unethical to infuse a placebo into chronically ill children, such as those with serious congenital heart disease or severe BPD, who would not personally benefit from the treatment. A total of 249 infants and young children were studied. Enrollees had either BPD (n 5 102), congenital heart disease (n 5 87), or prematurity alone (n 5 60). RSVIG was infused monthly (either 750 mg/kg, high dose; or 150 mg/kg, low dose); 89 control infants received no RSVIG. The treatment was well tolerated with mostly mild adverse reactions in about 3% of the 580 total infusions. One child had a serious reaction with respiratory failure 24 h later. This child had preexisting severe BPD and died of progressive Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest (17 patients treated, 18 given placebo). The IVIG treatments were well tolerated, and short- and long-term follow-up found the treatment to be safe. The treated patients had significant reductions in nasal virus (P , 0.01) and significant improvement in oxygenation 24 h after infusion (P , 0.05). However, the mean duration of hospitalization was not significantly reduced (3.94 days for 18 control patients and 3.06 days for 17 treated patients) (71). Further studies of parenteral IVIG therapy of RSV infections have not been done. Meissner et al. (92) conducted a prospective randomized, controlled trial of IVIG (Gammimune N; Cutter Biological, Miles, Inc., Berkeley, Calif.; NT titer, 950) to evaluate protection of 49 children with congenital heart disease or bronchopulmonary dysplasia (BPD) from RSV infection. Twenty-five patients received monthly infusions of 500 mg of IVIG per kg of body weight. Six RSV infections were observed in each group. Though the differences were not significant, the IVIG recipients tended to become less ill and were hospitalized for fewer days. The infusions were well tolerated and caused no evident morbidity. However, the IVIG lot used had insufficient NT titers to achieve protective levels (116) in the sera of the enrolled infants. Groothuis and colleagues reported that RSV infections were devastating for children with BPD (53). Studies were designed to test whether high-risk children (BPD, symptomatic congenital heart disease, or prematurity) could be protected from RSV infection or have less severe disease if passively immunized with IVIG during the RSV season. Multiple lots of several U.S.-licensed IVIG preparations were screened, but lots with NT titers exceeding 1,300 at a 5% concentration were not found. In a compromise, the tolerability and safety phase of the RSV prophylaxis trial was performed with a standard IVIG lot containing a suboptimal NT titer. Cotton rat studies had suggested that optimal protection required trough NT titers to be maintained in the range of 200 to 350 (114). The safety and tolerability portion of the trial was done with two lots of Gammimune-N. The geometric mean RSV NT titers at a 5% concentration were, respectively, 1,125 and 1,075 (54). Enrollees received their respective doses (one group received 500 mg/kg, one group received 600 mg/kg, and one group received 750 mg/kg) monthly from December through March of 1988 and 1989. Upon the safe completion of the pilot trial, the investigators expected to proceed to an efficacy trial of RSV immunoprophylaxis. Again, multiple lots of standard IVIG were screened. None had a sufficient NT titer to achieve adequate 30-day postinfusion antibody levels (trough titers) following sensibly sized, monthly IVIG infusions. CLIN. MICROBIOL. REV. VOL. 8, 1995 HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS 27 Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest FIG. 1. Relationship between serum RSV NT titer (complement-enhanced plaque reduction) at the time of RSV challenge and RSV concentrations (log10 PFU per gram; A/Long) recovered from lung homogenates (A) or from nasal turbinate homogenates (B). E, nonimmune control animals; 1, treated animals with RSV NT titers. Reproduced from reference 124 with permission of the publisher. respiratory failure 3 months later. Additionally, five other children with congenital heart disease died. Of the six deaths, three were in the high-dose and three were in the low-dose group. Three deaths were proximately associated with open heart surgery, and no death appeared related to the immunoglobulin infusions or to RSV infection. During the 3 years of the study, a total of 64 RSV infections were observed in the study population (19 in the high-dose group, 16 in the low-dose group, and 29 in the control group). Significant differences between the high-dose group and the control group, respectively, included fewer RSV respiratory tract infections (P 5 0.01), fewer hospitalizations (P 5 0.02), fewer hospital days (P 5 0.02), fewer days in the intensive care unit (P 5 0.05), and less use of ribavirin in the high-dose group (P 5 0.05) (55). The study demonstrates that passive immunization of selected high-risk infants and children with RSVIG during the RSV season substantially reduces RSV infection rates and the numbers of such children who require hospitalization for RSV 28 HEMMING ET AL. CLIN. MICROBIOL. REV. bronchiolitis and bronchopneumonia. RSV immunoprophylaxis remains investigational pending licensure review by the U.S. Food and Drug Administration. Three additional RSVIG trials are presently under way or just completed. One is a blinded, placebo-controlled, randomized treatment trial in children, without risk factors, hospitalized with RSV LRI. The second is a blinded, placebo-controlled, randomized treatment trial of high-risk children, such as those enrolled in the completed prophylaxis trial (55), who are hospitalized with RSV LRI. The third is a blinded, controlled prophylaxis trial further examining the safety of RSVIG prophylaxis in infants and young children with congen- FIG. 3. Yield (log10 PFU per gram) of RSV (A/Long) from the lungs of groups of cotton rats treated 1 day before RSV inoculation with 0.5 or 5.0 g of standard IVIG or RSVIG per kg of body weight. GM, geometric mean; NS, not significant. Reproduced from reference 124 with permission of the publisher. ital heart disease. The prevention or treatment of RSV infections with immunoglobulins remains investigational pending analysis of completed studies and review by the U.S. Food and Drug Administration. TOPICAL OR INHALED HUMAN IG AND RSVIG STUDIES IVIG Topical Studies in Cotton Rats and Owl Monkeys Pulmonary RSV infections in cotton rats (117), in owl monkeys (67), and in children may be treated by the parenteral FIG. 4. Yield (log10 PFU per gram) of RSV (A/Long) from the nasal turbinates of groups of cotton rats treated 1 day before RSV inoculation with 0.5 or 5.0 g of human IVIG or RSVIG per kg of body weight. GM, geometric mean; NS, not significant. Reproduced from reference 124 with permission of the publisher. Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest FIG. 2. Circulating RSV (A/Long) NT antibody titers (titer21) in groups of cotton rats 24 h following inoculation with 0.5 g of standard IVIG or RSVIG per kg of body weight compared with inoculation of 5.0 g of IG or RSVIG per kg. PRN, plaque reduction neutralization; NS, not significant. Reproduced from reference 124 with permission of the publisher. VOL. 8, 1995 HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS 29 FIG. 5. Comparison of the yield (log10 PFU per gram) of RSV (A/Long) from the lungs of cotton rats treated 3 days after infection with systemic (intraperitoneal) or topical (intranasal) human IG in various doses. Reprinted from reference 67 with permission of the publisher. injection of polyclonal human immune globulin (IG) containing sufficient amounts of NT antibody. Therefore, we wondered whether the topical administration of IG into the airway administered dropwise or by small-droplet aerosol, and containing known RSV NT antibody levels, might also be effective in the treatment of RSV LRI. Cotton rats were infected with RSV. At the height of infection, the rats were anesthetized and then treated with various doses of IG (Sandoglobulin, lot 2.370.069.0; RSV NT titer, 2,905) instilled intranasally in a volume of 0.1 ml per animal or inoculated intraperitoneally. Twenty-four hours after treatment, the animals were euthanized, their lungs and nasal tissues were homogenized, and RSV titers were determined on the homogenates by plaque assay on HEp-2 cells. Figure 5 depicts the comparative results of these experiments (67). The topical treatment, approximately 100 mg of IG per kg of body weight instilled dropwise intranasally in infected animals on day 3 of RSV infection, effected more than a 100-fold reduction of RSV in infected animals. Topical therapy required much less IG than parenteral therapy for a comparable response. This study also demonstrated that the effect did not come from in vitro neutralization. It was not associated with increased pulmonary pathology and did not prolong RSV infection (112, 113). RSV-infected owl monkeys were also treated by intratracheal instillation of IG at doses substantially below those necessary for comparable reductions of pulmonary RSV when treated by the parenteral route (70). In a follow-up study, it was also demonstrated that IG could be nebulized and delivered as a small-droplet aerosol for the treatment of RSV infections in cotton rats (109). The effects observed following the topical treatment of RSV infections were dose related. That is, pulmonary RSV titers correlated inversely with the amount of IG instilled or nebulized and also with the RSV NT titer of the IG. RSVIG Topical Studies in Cotton Rats Once RSVIG was available, a simple correlative study, using previously reported methods (109, 112), was performed to compare the therapeutic efficacies of RSVIG and standard IG (68). Groups of 12 adult cotton rats were infected with RSV. On day 3 after infection, the groups were treated with IG (NT titer, 1,100) or RSVIG (NT titer, 5,000) (0.05 ml per nostril) in either 5, 1, or 0.2% concentrations. The results of these experiments are depicted in Fig. 6 and 7. The intranasal treatment with IG or with RSVIG induced no reduction in the titers of RSV from the turbinates of any cotton rats in the treated groups, a result consistent with previous findings. In striking contrast, significant reductions in pulmonary viral titers (P , 0.001) were observed in both IG- and RSVIG-treated rats at 5% concentrations. However, the IG reduction was nearly 30-fold less than that observed with RSVIG. These studies again confirm the effectiveness of topical IG therapy in the FIG. 7. Comparison of the yield (log10 PFU per gram) of RSV (A/Long) from the lungs of cotton rats (12 animals per group) treated intranasally 3 days after infection (Long strain) with 0.1 ml of either 5.0 (■), 1.0 (s), or 0.2% (h) concentrations of human IG (Sandoglobulin) or RSVIG (lot 6, virally inactivated). Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest FIG. 6. Comparison of the yield (log10 PFU per gram) of RSV (A/Long) from the nasal turbinates of cotton rats (12 animals per group) treated intranasally 3 days after RSV infection with 0.1 ml of either 5.0 (■), 1.0 (s), or 0.2% (h) concentrations of human IG (Sandoglobulin) or RSVIG (lot 6, virally inactivated). 30 HEMMING ET AL. cotton rat (68). They also demonstrate the effectiveness of higher NT titers which, if the treatment becomes clinically relevant in humans, would permit the dramatic reduction in the amounts of antibody that might be required to effect a predicted outcome. RSV AND MONOCLONAL ANTIBODIES CONCLUSIONS Nearly 40 years have elapsed since the recovery and characterization of RSV. A large medical and virological literature attests that RSV infections are frequent causes of mild to severe respiratory illnesses often requiring hospitalization and occasionally responsible for death in infancy and early childhood in the United States and throughout the world. Primary and repeat infections fail in most cases to provide long-lasting immunity and so humans experience repeat RSV infections throughout life. Fortunately, most repeat infections in normal hosts are restricted to the upper respiratory tract. Humoral NT antibody protects the lung more efficiently than the upper airway and conjunctiva. This likely accounts for the type of reinfections with RSV. In this review, we have discussed clinical and laboratory data demonstrating that the maintenance of circulating NT antibodies above a critical level makes many high-risk patients significantly less likely to experience serious RSV LRI if infected. It would be preferable to maintain the requisite antibody levels by immunization. However, an effective vaccine is not yet available. In the absence of a vaccine, we have demonstrated that passive immunization of patients at high risk for a morbid outcome is a safe and effective alternative once RSVIG is available. Furthermore, animal studies suggest that there may be therapeutic indications for the use of hyperimmune RSV immunoglobulins for the parenteral and perhaps inhalation therapy of RSV infections in selected patients. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. REFERENCES 1. Abman, S. H., J. W. Ogle, N. Butler-Simon, C. M. Rumack, and F. J. Accurso. 1988. Role of respiratory syncytial virus in early hospitalizations for respiratory distress of young infants with cystic fibrosis. J. Pediatr. 113: 826–830. 2. Agius, G., G. Dindinaud, R. J. Biggar, R. Peyre, V. Vaillant, S. Ranger, J. Y. Poupet, M. F. Cisse, and M. Castets. 1990. An epidemic of respiratory syncytial virus in elderly people: clinical and serological findings. J. Med. Virol. 30:117–127. 3. Anderson, L. J., P. Bingham, and J. C. Hierholzer. 1988. Neutralization of respiratory syncytial virus by individual and mixtures of F and G protein monoclonal antibodies. J. Virol. 62:4232–4238. 4. Anderson, L. J., R. M. Hendry, L. T. Pierik, C. Tsou, and K. McIntosh. 1991. Multicenter study of strains of respiratory syncytial virus. J. Infect. Dis. 163:687–692. 5. Anderson, L. J., J. C. Hierholzer, P. G. Gingham, and Y. O. Stone. 1985. Microneutralization test for respiratory syncytial virus based on an enzyme immunoassay. J. Clin. Microbiol. 22:1050–1052. 6. Anderson, L. J., J. C. Hierholzer, Y. O. Stone, C. Tsou, and B. F. Fernie. 1986. Identification of epitopes on respiratory syncytial virus proteins by competitive binding immunoassay. J. Clin. Microbiol. 23:475–480. 7. Anderson, L. J., J. C. Hierholzer, C. Tsou, R. M. Hendry, B. F. Fernie, Y. 26. 27. 28. 29. 30. 31. Stone, and K. McIntosh. 1985. Antigenic characterization of respiratory syncytial virus strains with monoclonal antibodies. J. Infect. Dis. 151: 626–633. Beeler, J. A., and K. van Wyke Coelingh. 1989. Neutralization epitopes of the F glycoprotein of respiratory syncytial virus: effect of mutation on fusion function. J. Virol. 63:2941–2950. Beem, M. 1967. Repeated infections with respiratory syncytial virus. J. Immunol. 98:1115–1122. Beem, M., R. Egerer, and J. Anderson. 1964. Respiratory syncytial virus neutralizing antibodies in persons residing in Chicago, Illinois. Pediatrics 34:761–770. Beem, M., R. H. Wright, D. Hamre, R. Egerer, and M. Oehme. 1960. Association of the chimpanzee coryza agent with acute respiratory disease in children. N. Engl. J. Med. 263:523–530. Belshe, R. B., E. L. Anderson, and E. E. Walsh. 1993. Immunogenicity of purified F glycoprotein of respiratory syncytial virus: clinical and immune responses to subsequent natural infection in children. J. Infect. Dis. 168: 1024–1029. Belshe, R. B., L. S. Richardson, W. T. London, D. T. Slye, J. H. Lordeld, E. Camargo, D. A. Prevar, and R. M. Chanock. 1977. Experimental respiratory syncytial virus infection of four species of primates. J. Med. Virol. 1: 157–162. Belshe, R. B., L. P. Van Voris, and M. A. Mufson. 1982. Parenteral administration of live respiratory syncytial virus vaccine: results of a field trial. J. Infect. Dis. 145:311–319. Berkovich, S., and L. Taranko. 1964. Acute respiratory illness in the premature nursery associated with respiratory syncytial virus infections. Pediatrics 34:753–760. Brideau, R. J., R. R. Walters, M. A. Stier, and M. W. Wathen. 1989. Protection of cotton rats against human respiratory syncytial virus by vaccination with a novel chimeric FG glycoprotein. J. Gen. Virol. 70: 2637–2644. Bruhn, F. W., and A. S. Yeager. 1977. Respiratory syncytial virus in early infancy. Am. J. Dis. Child. 131:145–148. Buynak, E. B., R. E. Weibel, A. A. McLean, and M. R. Hilleman. 1978. Live respiratory syncytial virus vaccine administered parenterally. Proc. Soc. Exp. Biol. Med. 157:636–642. Cane, P. A., and C. R. Pringle. 1991. Respiratory syncytial virus heterogeneity during an epidemic: analysis by limited nucleotide sequencing (SH gene) and restriction mapping (N gene). J. Gen. Virol. 72:349–357. Chandwani, S., W. Borkowsky, K. Krasinski, R. Lawrence, and R. Welliver. 1990. Respiratory syncytial virus infection in human immunodeficiency virus-infected children. J. Pediatr. 117:251–254. Chanock, R., and L. Finberg. 1957. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). II. Epidemiologic aspects of infection in infants and young children. Am. J. Hyg. 66: 291–300. Chanock, R., B. Roizman, and R. Myers. 1957. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). I. Isolation, properties and characterization. Am. J. Hyg. 66:281–290. Chanock, R. M., H. W. Kim, A. J. Vargosko, A. DeLeva, K. M. Johnson, C. Cumming, and R. H. Parrott. 1961. Respiratory syncytial virus: I. Virus recovery and other observations during 1960 outbreak of bronchiolitis, pneumonia, and minor respiratory diseases in children. JAMA 176:647– 657. Chanock, R. M., and R. H. Parrott. 1965. Acute respiratory disease in infancy and childhood: present understanding and prospects for prevention. Pediatrics 36:21–39. Chanock, R. M., R. H. Parrott, A. Z. Kapikian, H. W. Kim, and C. D. Brandt. 1968. Possible role of immunological factors in pathogenesis of RS virus lower respiratory tract disease. Perspect. Virol. 6:125–140. Chanock, R. M., L. S. Richardson, R. B. Belshe, H. W. Kim, and R. M. Parrott. 1977. Prospects for prevention of bronchiolitis caused by respiratory syncytial virus. Pediatr. Res. 11:264–267. Chanock, R. M., C. B. Smith, W. T. Griedewald, R. H. Parrott, B. R. Forsyth, H. V. Coates, A. Z. Kapikian, and M. A. Gharpure. 1967. Resistance to parainfluenza and respiratory syncytial virus infection—implications for effective immunization and preliminary study of an attenuated strain of respiratory syncytial virus. Pan. Am. Health Organ. Sci. Publ. 147: 53–61. Chin, J., R. L. Magoffin, L. A. Shearer, J. H. Schieble, and E. H. Lennette. 1969. Field evaluation of a respiratory syncytial virus vaccine and a trivalent parainfluenza virus vaccine in a pediatric population. Am. J. Epidemiol. 89: 449–463. Coates, H. V., D. W. Alling, and R. M. Chanock. 1966. An antigenic analysis of respiratory syncytial virus isolates by a plaque reduction neutralization test. Am. J. Epidemiol. 83:299–313. Coates, H. V., L. Kendrick, and R. M. Chanock. 1963. Antigenic differences between two strains of respiratory syncytial virus. Proc. Soc. Exp. Biol. Med. 112:958–964. Collins, P. L., and G. M. Wertz. 1985. Gene products and genome organization of human respiratory syncytial (RS) virus, p. 297–310. In R. A. Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest Mouse, humanized mouse, and human monoclonal antibodies with RSV NT activity have been prepared and tested for prophylactic and therapeutic effects in RSV-infected mice and cotton rats (32, 34, 67, 134, 135). A review of RSV monoclonal antibodies is beyond the scope of this review. However, in the future, the use of recombinant DNA and monoclonal antibody technologies may permit the replacement of polyclonal human anti-RSV antibody preparations with highly specific monoclonal antibodies for prevention and treatment of human RSV infections. CLIN. MICROBIOL. REV. VOL. 8, 1995 32. 33. 34. 35. 36. 38. 39. 40. 41. 42. 43. 44. 45. 46. 47. 48. 49. 50. 51. 52. 53. 54. Lerner, R. M. Chanock, and F. Brown (ed.), Modern approaches to vaccines. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. Cote, P. J., Jr., B. F. Fernie, E. C. Ford, J. Wai-Kuo, and J. L. Gerin. 1981. Monoclonal antibodies to respiratory syncytial virus: detection of virus neutralization and other antigen-antibody systems using infected human and murine cells. J. Virol. Methods 3:137–147. Cristina, J., J. A. Lopez, C. Albo, B. Garcia-Barreno, J. Garcia, J. A. Melero, and A. Portela. 1990. Analysis of genetic variability in human respiratory syncytial virus by Rnase A mismatch cleavage method: subtype divergence and heterogeneity. Virology 174:126–134. Crowe, J. E., B. R. Murphy, R. M. Chanock, R. A. Williamson, C. F. Barbas III, and D. R. Burton. 1994. Recombinant human respiratory syncytial virus (RSV) monoclonal antibody Fab is effective therapeutically when introduced directly into the lungs of RSV-infected mice. Proc. Natl. Acad. Sci. USA 91:1386–1390. Cunningham, C. K., J. A. McMillan, and S. J. Gross. 1991. Rehospitalization for respiratory illness in infants of less than 32 weeks gestation. Pediatrics 88:527–532. Dreizin, R. S., L. O. Vyshnevetskaia, E. E. Bagdamian, O. D. Yankevich, L. B. Tarasova, and A. V. Klenova. 1971. Study of experimental respiratory syncytial virus infection in cotton rats: virologic and immunofluorescent studies. Vopr. Virusol. 16:670–676. (In Russian.) Englund, J. A., L. J. Anderson, and F. S. Rhame. 1991. Nosocomial transmission of respiratory syncytial virus in immunocompromised adults. J. Clin. Microbiol. 29:115–119. Englund, J. A., C. J. Sullivan, M. C. Jordan, L. P. Dehner, G. M. Vercellotti, and H. H. Balfour, Jr. 1988. Respiratory syncytial virus infection in immunocompromised adults. Ann. Intern. Med. 109:203–208. Fernie, G. F., and J. L. Gerin. 1982. Immunochemical identification of viral and nonviral proteins of the respiratory syncytial virus virion. Infect. Immun. 37:243–249. Fouillard, L., L. Mouthon, J. P. Laporte, F. Isnard, J. Stachowiak, M. Aoudjhane, J. C. Lucet, M. Wolf, F. Bricourt, L. Douay, M. Lopez, C. Marche, A. Najman, and N. C. Gorin. 1992. Severe respiratory syncytial virus pneumonia after autologous bone marrow transplantation: a report of three cases and a review. Bone Marrow Transplant. 9:97–100. Freymuth, F., J. Petitjean, P. Pothier, J. Brouard, and E. Norrby. 1991. Prevalence of respiratory syncytial virus subgroups A and B in France from 1982 to 1990. J. Clin. Microbiol. 29:653–655. Fulginiti, V. A., J. H. Arthur, D. S. Pearlman, and C. H. Kempe. 1968. Altered reactivity to measles virus: local reactions following attenuated measles virus immunization in children who previously received a combination of inactivated and attenuated vaccines. Am. J. Dis. Child. 115:671– 676. Fulginiti, V. A., J. J. Eller, A. W. Downie, and C. H. Kempe. 1967. Altered reactivity to measles virus: atypical measles in children previously immunized with inactivated measles vaccines. JAMA 202:1075–1080. Fulginiti, V. A., J. J. Eller, O. F. Sieber, J. W. Joyner, M. Minamitani, and G. Meiklejohn. 1969. Respiratory virus immunization. I. A field trial of two inactivated respiratory syncytial virus vaccines; an aqueous trivalent parainfluenza virus vaccine and an alum-precipitated respiratory syncytial virus vaccine. Am. J. Epidemiol. 89:435–448. Garcia-Barreno, B., C. Palomo, C. Penas, T. Delgado, P. Perez-Brena, and J. A. Melero. 1989. Marked differences in the antigenic structure of respiratory syncytial virus F and G glycoproteins. J. Virol. 63:925–932. Gardner, P. S., F. M. Elderkin, and A. H. Wall. 1964. Serological study of respiratory syncytial virus infections in infancy and childhood. Br. Med. J. 2:1570–1573. Gimenez, H. B., P. Cash, and W. T. Melvin. 1984. Monoclonal antibodies to human respiratory syncytial virus and their use in comparison of different virus isolates. J. Gen. Virol. 65:963–971. Gimenez, H. B., N. Hardman, H. M. Keir, and P. Cash. 1986. Antigenic variation between human respiratory syncytial virus isolates. J. Gen. Virol. 67:863–870. Glezen, W. P., A. Paredes, J. E. Allison, L. H. Taber, and A. L. Frank. 1981. Risk of respiratory syncytial virus infection for infants from low-income families in relationship to age, sex, ethnic group, and maternal antibody level. J. Pediatr. 98:708–715. Glezen, W. P., L. H. Taber, A. L. Frank, and J. A. Kasel. 1986. Risk of primary infection and reinfection with respiratory syncytial virus. Am. J. Dis. Child. 140:543–546. Graham, B. S., L. A. Bunton, J. Rowland, P. F. Wright, and D. T. Karzon. 1991. Respiratory syncytial virus infection in anti-m-treated mice. J. Virol. 65:4936–4942. Graham, B. S., T. H. Davis, Y.-W. Tang, and W. C. Gruber. 1993. Immunoprophylaxis and immunotherapy of respiratory syncytial virusinfected mice with respiratory syncytial virus-specific immune serum. Pediatr. Res. 34:167–172. Groothuis, J. R., K. M. Gutierrez, and B. A. Lauer. 1988. Respiratory syncytial virus infection in children with bronchopulmonary dysplasia. Pediatrics 82:199–203. Groothuis, J. R., M. J. Levin, W. Rodriguez, C. B. Hall, C. E. Long, H. W. 55. 56. 57. 58. 59. 60. 61. 62. 63. 64. 65. 66. 67. 68. 69. 70. 71. 72. 73. 74. 75. 76. 31 Kim, B. A. Lauer, and V. G. Hemming. 1991. Use of intravenous gamma globulin to passively immunize high-risk children against respiratory syncytial virus: safety and pharmacokinetics. Antimicrob. Agents Chemother. 35:1469–1473. Groothuis, J. R., E. A. F. Simoes, J. J. Levin, C. B. Hall, C. E. Long, W. J. Rodriguez, J. Arrobio, H. C. Meissner, D. R. Fulton, R. C. Welliver, D. A. Tristram, G. R. Siber, G. A. Prince, M. Van Raden, and V. G. Hemming. 1993. Prophylactic administration of respiratory syncytial virus immune globulin to high-risk infants and young children. N. Engl. J. Med. 329:1524– 1530. Gruber, W. C., S. Z. Wilson, B. J. Throop, and P. R. Wyde. 1987. Immunoglobulin administration and ribavirin therapy: efficacy in respiratory syncytial virus infection of the cotton rat. Pediatr. Res. 21:270–274. Hall, C. B., and R. G. Douglas, Jr. 1976. Respiratory syncytial virus and influenza: practical community surveillance. Am. J. Dis. Child. 130:615– 620. Hall, C. B., J. M. Geiman, R. G. Douglas, Jr., and M. P. Meagher. 1978. Control of nosocomial respiratory syncytial viral infections. Pediatrics 62: 728–732. Hall, C. B., A. E. Kopelman, R. G. Douglas, Jr., J. M. Geiman, and M. P. Meagher. 1979. Neonatal respiratory syncytial virus infection. N. Engl. J. Med. 300:393–396. Hall, C. B., K. R. Powell, N. E. MacDonald, C. L. Gala, M. E. Menegus, S. C. Suffin, and H. J. Cohen. 1986. Respiratory syncytial virus infection in children with compromised immune function. N. Engl. J. Med. 315:77–81. Hall, C. B., E. E. Walsh, C. E. Long, and K. C. Schnabel. 1991. Immunity to and frequency of reinfection with respiratory syncytial virus. J. Infect. Dis. 163:693–698. Hall, C. B., E. E. Walsh, K. C. Schnabel, C. E. Long, K. M. McConnochie, S. W. Hildreth, and L. J. Anderson. 1990. Occurrence of groups A and B of respiratory syncytial virus over 15 years: associated epidemiologic and clinical characteristics in hospitalized and ambulatory children. J. Infect. Dis. 162:1283–1290. Hamparian, V. V., A. Ketler, M. R. Hilleman, C. M. Reilly, L. McClelland, D. Cornfield, and J. Stokes, Jr. 1961. Studies of acute respiratory illness caused by respiratory syncytial virus. 1. Laboratory findings in 109 cases. Proc. Soc. Exp. Biol. Med. 106:717–722. Hamre, D., J. C. Lashof, J. A. Marshall, J. E. Cassidy, M. Smith, and C. R. Bennett. 1961. A study of viruses implicated in acute respiratory disease in Chicago from October, 1958, to June 1959. Am. Rev. Respir. Dis. 83:68–72. Harrington, R. D., T. M. Hooton, R. C. Hackman, G. A. Storch, B. Osborne, C. A. Gleaves, A. Benson, and J. D. Meyers. 1992. An outbreak of respiratory syncytial virus in a bone marrow transplant center. J. Infect. Dis. 165:987–993. Hemming, V. G., G. W. Fischer, and G. A. Prince. 1985. Giant cell pneumonia due to respiratory syncytial virus. Arch. Pathol. Lab. Med. 109: 213. (Letter.) Hemming, V. G., and G. A. Prince. 1992. Respiratory syncytial virus: babies and antibodies. Infect. Agents Dis. 1:24–32. Hemming, V. G., and G. A. Prince. Unpublished data. Hemming, V. G., G. A. Prince, R. L. Horswood, W. T. London, B. R. Murphy, E. E. Walsh, G. W. Fischer, L. E. Weisman, P. A. Baron, and R. M. Chanock. 1985. Studies of passive immunotherapy for infections of respiratory syncytial virus in the respiratory tract of a primate model. J. Infect. Dis. 152:1083–1087. Hemming, V. G., G. A. Prince, W. T. London, P. A. Baron, R. Brown, and R. M. Chanock. 1988. Topically administered immunoglobulin reduces pulmonary respiratory syncytial virus shedding in owl monkeys. Antimicrob. Agents Chemother. 32:1269–1270. Hemming, V. G., W. Rodriguez, H. W. Kim, C. D. Brandt, R. H. Parrott, B. Burch, G. A. Prince, P. A. Baron, R. J. Fink, and G. Reaman. 1987. Intravenous immunoglobulin treatment of respiratory syncytial virus infections in infants and young children. Antimicrob. Agents Chemother. 31: 1882–1886. Henderson, F. W., A. M. Collier, W. A. Clyde, and F. W. Denny. 1979. Respiratory-syncytial-virus infections, reinfections and immunity: a prospective, longitudinal study in young children. N. Engl. J. Med. 300:530– 534. Hertz, M. I., J. A. Englund, D. Snover, P. B. Bitterman, and P. B. McGlave. 1989. Respiratory syncytial virus-induced acute lung injury in adult patients with bone marrow transplants: a clinical approach and review of the literature. Medicine (Baltimore) 68:269–281. Hierholzer, J. C., and M. S. Hirsch. 1979. Croup and pneumonia in human infants associated with a new strain of respiratory syncytial virus. J. Infect. Dis. 140:826–828. Homa, F. L., R. J. Brideau, D. J. Lehman, D. R. Thomsen, R. A. Olmsted, and M. W. Wathen. 1993. Development of a novel subunit vaccine that protects cotton rats against both human respiratory syncytial virus and human parainfluenza virus type 3. J. Gen. Virol. 74:1995–1999. Johnson, K. M., H. H. Bloom, M. A. Mufson, and R. M. Chanock. 1962. Natural reinfection of adults by respiratory syncytial virus: possible relation to mild upper respiratory disease. N. Engl. J. Med. 267:68–72. Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest 37. HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS 32 HEMMING ET AL. from lungs with virus-specific cytotoxic T cells. J. Virol. 65:4494–4497. 101. Murphy, B. R., B. S. Graham, G. A. Prince, E. E. Walsh, R. M. Chanock, D. T. Karzon, and P. F. Wright. 1986. Serum and nasal-wash immunoglobulin G and A antibody response of infants and children to respiratory syncytial virus F and G glycoproteins following primary infection. J. Clin. Microbiol. 23:1009–1014. 102. Murphy, B. R., A. Sotnikov, P. R. Paradiso, S. W. Hildreth, A. B. Jenson, R. B. Baggs, L. Lawrence, J. J. Zubak, R. M. Chanock, J. A. Beeler, and G. A. Prince. 1989. Immunization of cotton rats with the fusion (F) and large (G) glycoproteins of respiratory syncytial virus (RSV) protects against RSV challenge without potentiating RSV disease. Vaccine 7:533–540. 103. Nadal, D., B. Albini, E. Schlapfer, C. Chen, L. Brodsky, and P. L. Ogra. 1991. Tissue distribution of mucosal antibody-producing cells specific for respiratory syncytial virus in severe combined immune deficiency (SCID) mice engrafted with human tonsils. Clin. Exp. Immunol. 85:358–364. 104. Nader, P. R., M. Horwitz, and J. Rousseau. 1968. Atypical exanthem following exposure to natural measles: 14 cases in children previously inoculated with killed vaccine. J. Pediatr. 72:22–28. 105. Navas, L., E. Wang, V. de Carvalho, and J. Robinson. 1992. Improved outcome of respiratory syncytial virus infection in a high-risk hospitalized population of Canadian children. J. Pediatr. 121:348–353. 106. Ogilvie, M. M., S. Vathenen, M. Radford, J. Codd, and S. Key. 1981. Maternal antibody and respiratory syncytial virus infection in infancy. J. Med. Virol. 7:263–271. 107. Olmsted, R. A., R. M. L. Buller, B. R. Murphy, J. A. Beeler, P. L. Collins, and W. T. London. 1988. Evaluation in nonhuman primates of the safety, immunogenicity, and efficacy of recombinant vaccinia viruses expressing the F or G glycoprotein of respiratory syncytial virus, p. 205–210. In R. M. Chanock, R. A. Lerner, F. Brown, and H. Ginsberg (ed.), Vaccines 88: modern approaches to new vaccines including prevention of AIDS. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 108. Parrott, R. H., A. J. Vargosko, H. W. Kim, C. Cumming, H. Turner, R. J. Huebner, and R. M. Chanock. 1961. Respiratory syncytial virus: II. Serologic studies over a 34-month period of children with bronchiolitis, pneumonia, and minor respiratory diseases. JAMA 176:653–657. 109. Piazza, F. M., S. A. Johnson, M. G. Ottolini, H. J. Schmidt, M. E. R. Darnall, V. G. Hemming, and G. A. Prince. 1992. Immunotherapy of respiratory syncytial virus infection in cotton rats (Sigmodon fulviventer) using IgG in a small-particle aerosol. J. Infect. Dis. 166:1422–1424. 110. Porter, D. D., K. B. Muck, and G. A. Prince. 1980. The age dependence of respiratory syncytial virus growth in ferret lung can be shown in organ monolayer cultures. Clin. Immunol. Immunopathol. 15:415–423. 111. Prince, G. A. 1975. The pathogenesis of respiratory syncytial virus infection in infant ferrets. Doctoral dissertation. University of California at Los Angeles. 112. Prince, G. A., V. G. Hemming, R. L. Horswood, P. A. Baron, and R. M. Chanock. 1987. Effectiveness of topically administered neutralizing antibodies in experimental immunotherapy of respiratory syncytial virus infection in cotton rats. J. Virol. 61:1851–1854. 113. Prince, G. A., V. G. Hemming, R. L. Horswood, P. A. Baron, B. R. Murphy, and R. M. Chanock. 1990. Mechanism of antibody-mediated viral clearance in immunotherapy of respiratory syncytial virus infection in cotton rats. J. Virol. 64:3091–3092. 114. Prince, G. A., V. G. Hemming, R. L. Horswood, and R. M. Chanock. 1985. Immunoprophylaxis and immunotherapy of respiratory syncytial virus infection in the cotton rat. Virus Res. 3:193–206. 115. Prince, G. A., R. L. Horswood, E. Camargo, D. Koenig, and R. M. Chanock. 1983. Mechanisms of immunity to respiratory syncytial virus in cotton rats. Infect. Immun. 42:81–87. 116. Prince, G. A., R. L. Horswood, and R. M. Chanock. 1985. Quantitative aspects of passive immunity to respiratory syncytial virus infection in infant cotton rats. J. Virol. 55:517–520. 117. Prince, G. A., A. B. Jenson, V. G. Hemming, B. R. Murphy, E. E. Walsh, R. L. Horswood, and R. M. Chanock. 1986. Enhancement of respiratory syncytial virus pulmonary pathology in cotton rats by prior intramuscular inoculation of formalin-inactivated virus. J. Virol. 57:721–728. 118. Prince, G. A., A. B. Jenson, R. L. Horswood, E. Camargo, and R. M. Chanock. 1978. The pathogenesis of respiratory syncytial virus infection in cotton rats. Am. J. Pathol. 93:771–792. 119. Prince, G. A., and D. D. Porter. 1976. The pathogenesis of respiratory syncytial virus in infant ferrets. Am. J. Pathol. 82:339–352. 120. Prince, G. A., S. C. Suffin, D. A. Prevar, E. Camargo, D. Sly, W. T. London, and R. M. Chanock. 1979. Respiratory syncytial virus infection in owl monkeys: viral shedding, immunological response, and associated illness caused by wild-type virus and two temperature-sensitive mutants. Infect. Immun. 26:1009–1013. 121. Reilly, C. M., J. Stokes, Jr., L. McClelland, D. Cornfield, V. V. Hamparian, A. Ketler, and M. R. Hilleman. 1961. Studies of acute respiratory illness caused by respiratory syncytial virus. 3. Clinical and laboratory findings. N. Engl. J. Med. 264:1176–1182. 122. Richardson, L. S., R. B. Belshe, W. T. London, D. L. Sly, D. A. Prevar, E. Camargo, and R. M. Chanock. 1978. Evaluation of five temperature- Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest 77. Johnson, P. R., Jr., R. A. Olmsted, G. A. Prince, B. R. Murphy, D. W. Alling, E. E. Walsh, and P. L. Collins. 1987. Antigenic relatedness between glycoproteins of human respiratory syncytial virus subgroups A and B: evaluation of the contributions of F and G glycoproteins to immunity. J. Virol. 61:3163–3166. 78. Johnson, R. A., G. A. Prince, S. C. Suffin, R. L. Horswood, and R. M. Chanock. 1982. Respiratory syncytial virus infection in cyclophosphamidetreated cotton rats. Infect. Immun. 37:369–373. 79. Kapikian, A. Z., R. H. Mitchell, R. M. Chanock, R. A. Shvedoff, and C. E. Stewart. 1969. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am. J. Epidemiol. 89:405–421. 80. Kim, H. W., J. O. Arrobio, C. D. Brandt, B. C. Jeffries, G. Pyles, J. L. Reid, R. M. Chanock, and R. H. Parrott. 1973. Epidemiology of respiratory syncytial virus infection in Washington, D.C. I. Importance of the virus in different respiratory tract syndromes and temporal distribution of infection. Am. J. Epidemiol. 98:216–225. 81. Kim, H. W., J. G. Canchola, C. D. Brandt, G. Pyles, R. M. Chanock, K. Jensen, and R. H. Parrott. 1969. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 89:422–434. 82. Kravetz, H. M., V. Knight, R. M. Chanock, J. A. Morris, K. M. Johnson, D. Rifkind, and J. P. Utz. 1961. Respiratory syncytial virus: III. Production of illness and clinical observations in adult volunteers. JAMA 176:657–667. 83. Kurlandsky, L. E., G. French, P. M. Webb, and D. D. Porter. 1988. Fatal respiratory syncytial virus pneumonitis in a previously healthy child. Am. Rev. Respir. Dis. 138:468–472. 84. Lamprecht, C. L., H. E. Krause, and M. A. Mufson. 1976. Role of maternal antibody in pneumonia and bronchiolitis due to respiratory syncytial virus. J. Infect. Dis. 134:211–217. 85. Law, B. J., and V. de Carvalho. 1993. Respiratory syncytial virus infections in hospitalized Canadian children: regional differences in patient populations and management practices. Pediatr. Infect. Dis. J. 12:659–663. 86. MacDonald, N. E., C. B. Hall, S. C. Suffin, C. Alexson, P. J. Harris, and J. A. Manning. 1982. Respiratory syncytial viral infections in infants with congenital heart disease. N. Engl. J. Med. 307:397–400. 87. McClelland, L., M. R. Hilleman, V. V. Hamparian, A. Ketler, C. M. Reilly, D. Cornfield, and J. Stokes. 1961. Studies of acute respiratory illnesses caused by respiratory syncytial virus. 2. Epidemiology and assessment of importance. N. Engl. J. Med. 264:1169–1175. 88. McConnochie, K. M., C. B. Hall, E. E. Walsh, and K. J. Roghmann. 1990. Variation in severity of respiratory syncytial virus infections with subtype. J. Pediatr. 117:52–62. 89. McIntosh, E. D. G., L. M. de Silva, and R. K. Oates. 1993. Clinical severity of respiratory syncytial virus group A and B infection in Sydney, Australia. Pediatr. Infect. Dis. J. 12:815–819. 90. McIntosh, K., H. B. Masters, I. Orr, R. K. Chao, and R. M. Barkin. 1978. The immunologic response to infection with respiratory syncytial virus in infants. J. Infect. Dis. 138:24–32. 91. McKay, E., P. Higgins, D. Tyrrell, and C. Pringle. 1988. Immunogenicity and pathogenicity of temperature-sensitive modified respiratory syncytial virus in adult volunteers. J. Med. Virol. 25:411–421. 92. Meissner, H. C., D. R. Fulton, J. R. Groothuis, R. L. Geggel, G. R. Marx, V. G. Hemming, T. Hougen, and D. R. Snydman. 1993. Controlled trial to evaluate protection of high-risk infants against respiratory syncytial virus disease by using standard intravenous immune globulin. Antimicrob. Agents Chemother. 37:1655–1658. 93. Moler, F. W., A. S. Khan, J. N. Meliones, J. R. Custer, J. Palmisano, and T. C. Shope. 1992. Respiratory syncytial virus morbidity and mortality estimates in congenital heart disease patients: a recent experience. Crit. Care Med. 20:1406–1413. 94. Monto, A. S., and S. K. Lim. 1971. The Tecumseh study of respiratory illness. III. Incidence and periodicity of respiratory syncytial virus and Mycoplasma pneumoniae infections. Am. J. Epidemiol. 94:290–302. 95. Monto, A. S., and S. Ohmit. 1990. Respiratory syncytial virus in a community population: circulation of subgroups A and B since 1965. J. Infect. Dis. 161:781–783. 96. Morris, J. A., R. E. Blount, and R. E. Savage. 1956. Recovery of cytopathogenic agent from chimpanzees with coryza. Proc. Soc. Exp. Biol. Med. 92: 544–549. 97. Muelenaer, P. M., F. W. Henderson, V. G. Hemming, E. E. Walsh, L. J. Anderson, G. A. Prince, and B. R. Murphy. 1991. Group-specific serum antibody responses in children with primary and recurrent respiratory syncytial virus infections. J. Infect. Dis. 164:15–21. 98. Mufson, M. A., B. Akerlind-Stopner, C. Orvell, R. B. Belshe, and E. Norrby. 1991. A single-season epidemic with respiratory syncytial virus subgroup B2 during 10 epidemic years, 1978 to 1988. J. Clin. Microbiol. 29:162–165. 99. Mufson, M. A., R. B. Belshe, C. Orvell, and E. Norrby. 1987. Subgroup characteristics of respiratory syncytial virus strains recovered from children with two consecutive infections. J. Clin. Microbiol. 25:1535–1539. 100. Munoz, J. L., C. A. McCarthy, M. E. Clark, and C. B. Hall. 1991. Respiratory syncytial virus infection in C57BL/6 mice: clearance of virus CLIN. MICROBIOL. REV. VOL. 8, 1995 123. 124. 125. 126. 127. 129. 130. 131. 132. 133. 134. 135. 136. 137. sensitive mutants of respiratory syncytial virus in primates. I. Viral shedding, immunologic response and associated illness. J. Med. Virol. 3:91–100. Richardson-Wyatt, L. S., R. B. Belshe, W. T. London, D. L. Sly, E. Camargo, and R. M. Chanock. 1981. Respiratory syncytial virus antibodies in nonhuman primates and domestic animals. Lab. Anim. Sci. 31:413–415. Siber, G. R., D. Leombruno, J. Leszczynski, J. McIver, D. Bodkin, R. Gonin, C. M. Thompson, E. E. Walsh, P. A. Piedra, V. G. Hemming, and G. A. Prince. 1994. Comparison of antibody concentrations and protective activity of respiratory syncytial virus immune globulin and conventional immune globulin. J. Infect. Dis. 169:1368–1373. Siber, G. R., J. Leszczynski, V. Pena-Cruz, C. Ferren-Gardner, R. Anderson, V. G. Hemming, E. E. Walsh, J. Burns, K. McIntosh, R. Gonin, and L. J. Anderson. 1992. Protective activity of a human respiratory syncytial virus immune globulin prepared from donors screened by microneutralization assay. J. Infect. Dis. 165:456–463. Siqueira, M. M., J. P. Nascimento, and L. J. Anderson. 1991. Antigenic characterization of respiratory syncytial virus group A and B isolates in Rio de Janeiro, Brazil. J. Clin. Microbiol. 29:557–559. Sriskandan, S., and S. Shaunak. 1993. Respiratory syncytial virus infection in an adult with AIDS. Clin. Infect. Dis. 17:1065. Storch, G. A., L. J. Anderson, C. S. Park, C. Tsou, and D. E. Dohner. 1991. Antigenic and genomic diversity within group A respiratory syncytial virus. J. Infect. Dis. 163:858–861. Stretton, M., S. J. Ajizian, I. Mitchell, and C. J. L. Newth. 1992. Intensive care course and outcome of patients infected with respiratory syncytial virus. Pediatr. Pulmonol. 13:143–150. Suffin, S. C., G. A. Prince, K. B. Muck, and D. D. Porter. 1979. Immunoprophylaxis of respiratory syncytial virus in the infant ferret. J. Immunol. 123:10–14. Sullender, W. M., M. A. Mufson, L. J. Anderson, and G. W. Wertz. 1991. Genetic diversity of the attachment protein of subgroup B respiratory syncytial viruses. J. Virol. 65:5425–5434. Suto, T., N. Yano, M. Ikeda, M. Miyamoto, S. Takai, S. Shigeta, Y. Hinuma, and N. Ishida. 1965. Respiratory syncytial virus infection and its serologic epidemiology. Am. J. Epidemiol. 82:211–224. Taylor, C. E., A. W. Craft, J. Kernahan, R. Millman, M. M. Reid, R. Scott, and G. L. Toms. 1990. Local antibody production and respiratory syncytial virus in children with leukaemia. J. Med. Virol. 30:277–281. Taylor, G., J. Furze, P. R. Tempest, P. Bremner, F. J. Carr, and W. J. Harris. 1991. Humanized monoclonal antibody to respiratory syncytial virus. Lancet 337:1411–1422. Taylor, G., E. J. Scott, M. Bew, B. F. Fernie, P. J. Cote, A. P. Collins, M. Hughes, and J. Jebbett. 1984. Monoclonal antibodies protect against respiratory syncytial virus infection in mice. Immunology 52:137–142. Taylor, G., E. J. Stott, M. Hughes, and A. P. Collins. 1984. Respiratory syncytial virus infection in mice. Infect. Immun. 43:649–655. Thomas, E., M. J. Margach, C. Orvell, B. Morrison, and E. Wilson. 1994. Respiratory syncytial virus subgroup B dominance during one winter season 33 between 1987 and 1992 in Vancouver, Canada. J. Clin. Microbiol. 32:238–242. 138. Tristram, D. A., R. C. Welliver, C. K. Mohar, D. A. Hogerman, S. W. Hildreth, and P. Paradiso. 1993. Immunogenicity and safety of respiratory syncytial virus subunit vaccine in seropositive children 18–36 months old. J. Infect. Dis. 167:191–195. 139. Tsutsumi, H., M. Onuma, K. Suga, T. Honjo, Y. Chiba, S. Chiba, and P. L. Ogra. 1988. Occurrence of respiratory syncytial virus subgroup A and B strains in Japan, 1980 to 1987. J. Clin. Microbiol. 26:1171–1174. 140. Walsh, E. E., M. W. Brandriss, and J. J. Schlesinger. 1985. Purification and characterization of the respiratory syncytial virus fusion protein. J. Gen. Virol. 66:409–415. 141. Walsh, E. E., C. B. Hall, M. Briselli, M. W. Brandriss, and J. J. Schlesinger. 1987. Immunization with glycoprotein subunits of respiratory syncytial virus to protect cotton rats against viral infection. J. Infect. Dis. 155:1198–1204. 142. Walsh, E. E., J. J. Schlesinger, and M. W. Brandriss. 1984. Purification and characterization of GP90, one of the envelope glycoproteins of respiratory syncytial virus. J. Gen. Virol. 65:761–767. 143. Ward, K. A., P. R. Lambden, M. M. Ogilvie, and P. J. Watt. 1983. Antibodies to respiratory syncytial virus polypeptides and their significance in human infection. J. Gen. Virol. 64:1867–1876. 144. Waris, M. 1991. Pattern of respiratory syncytial virus epidemics in Finland: two-year cycles with alternating prevalence of groups A and B. J. Infect. Dis. 163:464–469. 145. Wathen, M. W., R. J. Brideau, and D. R. Thomsen. 1989. Immunization of cotton rats with the human respiratory syncytial virus F glycoprotein produced using a baculovirus vector. J. Infect. Dis. 159:255–264. 146. Wathen, M. W., R. J. Brideau, D. R. Thomsen, and B. R. Murphy. 1989. Characterization of a novel human respiratory syncytial virus chimeric FG glycoprotein expressed using a baculovirus vector. J. Gen. Virol. 70:2625– 2635. 147. Wilson, E., C. Orvell, B. Morrison, and E. Thomas. 1990. Pediatric RSV infection during two winter seasons in British Columbia: a role for subgroup analysis in young children? Can. J. Infect. Dis. 1:112–116. 148. Wong, D. T., M. Rosenband, K. Hovey, and P. L. Ogra. 1985. Respiratory syncytial virus in immunosuppressed animals: implications in human infection. J. Med. Virol. 17:359–370. 149. Wright, P. F., R. B. Belshe, H. W. Kim, L. P. Van Voris, and R. M. Chanock. 1982. Administration of a highly attenuated, live respiratory syncytial virus vaccine to adults and children. Infect. Immun. 37:397–400. 150. Wright, P. F., T. Shinozaki, W. Fleet, S. H. Sell, J. Thompson, and D. T. Karzon. 1976. Evaluation of a live, attenuated respiratory syncytial virus vaccine in infants. J. Pediatr. 88:931–936. 151. Wulff, H., P. Kidd, and H. A. Wenner. 1964. Respiratory syncytial virus: observations on antigenic heterogeneity. Proc. Soc. Exp. Biol. Med. 115: 240–243. 152. Wyde, P. R., C.-S. Sun, and V. Knight. 1983. Replication of respiratory syncytial virus in immunodeficient mice. J. Reticulo-endothel. Soc. 34:125– 129. Downloaded from http://cmr.asm.org/ on September 9, 2014 by guest 128. HYPERIMMUNE GLOBULINS IN PREVENTION OF RSV INFECTIONS



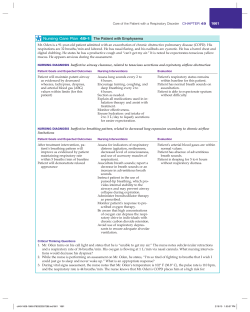






© Copyright 2026