
Mending the Broken Heart Guest Editorial Lead Article
Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Guest Editor: Michael R. Rosen, MD Mending the Broken Heart Guest Editorial Mending the broken heart - M. R. Rosen 3 Lead Article Regenerating the heart: new progress in gene/cell therapy to restore normal mechanical and electrical function - I. S. Cohen, G. R. Gaudette 7 Expert Answers to Three Key Questions Gene therapy for myocardial infarction–associated congestive heart failure: how far have we got? - H. K. Hammond, T. Tang 29 Does cell therapy for myocardial infarction and heart failure work? - K. C. Wollert, H. Drexler 37 Gene and cell therapy for life-threatening cardiac arrhythmias: will they replace drugs, surgery, and devices? - M. R. Rosen, P. Danilo Jr, R. B. Robinson 44 Fascinoma Cardiologica Matters @ Heart: Doppler and his principle - R. J. Bing 53 Summaries of Ten Seminal Papers - D. H. Lau 57 Evidence that human cardiac myocytes divide after myocardial infarction – A. P. Beltrami and others Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers – I. Potapova and others Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes Regenerating the heart – M. A. Laflamme and C. E. Murry I. Kehat and others Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction – D. L. Kraitchman and others Biological pacemaker created by gene transfer – J. Miake Molecular ablation of ventricular tachycardia after myocardial infarction – T. Sasano and others and others Heart regeneration in zebrafish – K. D. Poss and others Theoretical impact of the injection of material into the myocardium: a finite element model simulation – S. T. Wall Adult cardiac stem cells are multipotent and support myocardial regeneration – A. P. Beltrami and others and others Bibliography of One Hundred Key Papers 1 69 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Editors in Chief Ferrari R, MD, PhD Dept of Cardiology, Arcispedale S. Anna University of Ferrara, Ferrara, Italy Hearse DJ, BSc, PhD The Cardiothoracic Centre, The Rayne Institute St Thomas’ Hospital, London, UK Consulting Editors Avkiran M, PhD Cardiovascular Research The Rayne Institute St Thomas’ Hospital London, UK Bassand JP, MD Dept of Cardiology University Hospital Jean Minjoz Besançon, France Bertrand ME, MD Hôpital Cardiologique Lille, France Bolli R, MD Division of Cardiology University of Louisville Louisville, KY, USA Camm JA, MD Dept of Cardiac and Vascular Sciences St George’s University of London London, UK Coats A, MD Faculty of Medicine University of Sydney Sydney, Australia Cobbe SM, MD Dept of Medical Cardiology Glasgow Royal Infirmary Glasgow, UK Cohn JN, MD Rasmussen Center for Cardiovascular Disease Prevention Minneapolis, MN, USA Cokkinos DV, MD 1st Cardiology Dept Onassis Cardiac Surgery Center Athens, Greece Cowie M, MD, PhD Dept of Clinical Cardiology National Heart & Lung Institute London, UK Danchin N, MD Dept of Cardiology Hôpital Européen Georges Pompidou Paris, France Dargie HJ, MD Cardiac Research Western Infirmary Glasgow, UK Di Pasquale G, MD Dpt of Cardiology Maggiore Hospital Bologna, Italy Dzau VJ, MD Duke University Medical Center & Health System DUMC Durham, NC, USA Fernandez-Aviles F, MD Institute of Hematology and Oncology, IDIBAPS Hospital University Clinic of Barcelona Barcelona, Spain Fox KM, MD Dept of Cardiology Royal Brompton Hospital London, UK Fox KA, MD Dept of Cardiological Research University of Edinburgh Edinburgh, UK Fuster V, MD, PhD Cardiovascular Institute Mount Sinai Medical Center New York, NY, USA Hasenfuss G, MD Dept of Cardiology Georg-August Universität Göttingen, Germany Hori M, MD, PhD Dept of Internal Medicine and Therapeutics Osaka University Graduate School of Medicine Osaka, Japan Katz AM, MD University of Connecticut School of Medicine Farmington, CT, USA Komajda M, MD Dept of Cadiology CHU Pitié-Salpêtrière Paris, France Komuro I, MD, PhD Dept of Cardiovascular Sciences & Medicine Chiba University Graduate School of Medicine Chiba, Japan Lakatta EG, MD National Institute on Aging Gerontology Research Center Baltimore, MD, USA Libby P, MD Cardiovascular Medicine Brigham & Women’s Hospital Boston, MA, USA Lonn E, MD Hamilton Health Sciences General Site Hamilton, Ontario, Canada Lopez-Sendon JL, MD CCU Dept of Cardiology Hospital University Gregorio Maranon Madrid, Spain Maggioni AP, MD ANMC Research Center Firenze, Italy Marber MS, MD, PhD Cardiovascular Research The Rayne Institute St Thomas’ Hospital London, UK Sleight P, MD Dept of Cardiovascular Medicine John Radcliffe Hospital Oxford, UK Soler-Soler J, MD Dept of Cardiology Hospital General Vall d’Hebron Barcelona, Spain Oto A, MD Medical Office, Hacettepe University School of Medicine Ankara, Turkey Steg PG, MD Dept of Cardiology Hôpital Bichat–Claude Bernard Paris, France Patrono C, MD Dept of Pharmacology University La Sapienza Rome, Italy Swedberg K, MD, PhD Dept of Medicine Sahlgrenska University Hospital Ostra Göteborg, Sweden Pepine CJ, MD Dept of Medicine University of Florida Gainesville, FL, USA Rapezzi C, MD Institute of Cardiology University of Bologna Bologna, Italy Remme WJ, MD, PhD Sticares Foundation Rotterdam, The Netherlands Rosen MR, MD Dept of Pharmacology & Pediatrics Columbia University College of Physicians & Surgeons New York, NY, USA Ruzyllo W, MD National Institute of Cardiology Warsaw, Poland Ryden L, MD, PhD Dept of Cardiology Karolinska University Hospital Solna Stockholm, Sweden Schneider MD, MD Baylor College of Medicine Houston, TX, USA Seabra-Gomes RJ, MD Instituto do Coracao Hospital Santa Cruz Carnaxide, Portugal Sechtem U, MD Dept of Internal Medicine & Cardiology Robert Bosch Krankenhaus Stuttgart, Germany Simoons ML, MD Thoraxcenter Erasmus University Medical Center Rotterdam, The Netherlands 2 Tardif JC, MD Montreal Heart Institute Montreal, Quebec, Canada Tavazzi L, MD Division of Cardiology Policlinico San Matteo IRCCS Pavia, Italy Tendera M, MD 3rd Division of Cardiology Silesian School of Medicine Katowice, Poland Vanhoutte PM, MD Dept of Pharmacology University of Hong Kong Faculty of Medicine Hong Kong, China Widimsky P, MD, PhD Vinohrady Cardiocenter Charles University Hospital Prague, Czech Republic Wijns WC, MD Cardiovascular Center Aalst OLV Hospital, Aalst, Belgium Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Guest Editorial Michael R. Rosen, MD Center for Molecular Therapeutics - Columbia University - New York, NY - USA MENDING THE BROKEN HEART W hen David Hearse and Roberto Ferrari invited me to edit this issue and to contribute one of the articles I had mixed feelings. While the title proposed suggests a romantic paperback novel, the subject matter it incorporates is potentially groundbreaking: arguably, gene and cell therapies are fields that will apply to much of cardiovascular and other medical therapeutics in the future. The beginnings have been rocky, especially with regard to gene therapy. This largely reflects the toxicity of viral vectors used in the initial gene therapies that were administered to patients. Cell therapy has had a more successful beginning, first appearing on the scene in 1956. In that year, E. Donnall Thomas obtained long-term survival by transplanting bone marrow into a patient with leukemia. Since that time both the efficacy and safety of bone marrow transplant have been documented and detailed for the treatment of certain cancers and immunodeficiency diseases. Obviously there are toxicities and shortcomings, but the life-saving nature of the therapy is unquestionable. And this history has provided assurance to subsequent investigators studying marrowderived cells, assurance that the cells they deliver to patients likely will cause no harm. Important with regard to the safety issue is that in most instances the cells administered have been autologous. Given the limited safety concerns, there has been rapid advancement of cell therapy in humans with myocardial infarction and/or with heart failure. There has been almost marginal benefit, with the occasional report looking hopeful, but the safety of the procedure appears to have been validated. So if gene therapy has produced death and disease and cell therapy has been shown safe if not necessarily effective (except for bone marrow transplantation as stated above), why devote an issue to “Mending the Broken Heart?” My own prejudice is that ••• Michael R. Rosen, MD, Gustavus A. Pfeiffer Professor of Pharmacology, Professor of Pediatrics, Director, Center for Molecular Therapeutics, Columbia University, PH 7W-321, 630 West 168 Street, New York, NY, 10032, USA (e-mail: [email protected]) 3 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Guest Editorial - Michael R. Rosen, MD ••• gene and cell therapies are at a developmental stage not unlike antimicrobials in the 1930s. Sulfa drugs then appeared in a field that previously depended on folk medicine, carbolic acid, arsenicals, and the like. And although sulfa drugs were far from an ideal antimicrobial, they were not only a vast improvement, but they presaged the arrival of antibiotics in the clinic in the 1940s. To succeed, investigators had to accept the germ theory of disease, a nineteenth century concept verified scientifically by Koch in 1875, and then learn a new language: partly that of the germs themselves and partly that of the molds and other sources from which they made their antibiotics. Whereas Tyndall in 1875 had already noted the antimicrobial properties of Penicillium, it was not until 1928 that Fleming replicated the observation and did one essential additional experiment: he used the soup in which the mold grew to treat bacterial colonies, saw that it lysed them, called the soup penicillin and gave new impetus to a burgeoning field of therapy. All that remained was to develop a stable, clinically useful system to make penicillin, and this required another decade. Those individuals now working in the field of cell and gene therapy are also having to learn a language; arguably a far more obscure language than that of the bacilli and molds that dot the antibacterial landscape. What are the signals and factors that determine stem cell growth and fate? How can these be manipulated safely? How reproducibly can cells be made to grow, to mature, but not to evolve into neoplasms? The list is almost endless. These issues and others relating to the use of cells to repair/regenerate myocardium are considered in this issue by Ira Cohen and Glenn Gaudette. The segue from the cells under investigation and problems in understanding their biology to their use in clinical settings of myocardial infarction and heart failure is considered by Kai Wollert and Helmut Drexler. Review of the types of cells available, the means for their administration and the clinical successes and limitations to date is both encouraging and chastening as it indicates how far we have to go to understand what we are doing in the clinic and how to do it better. Kirk Hammond and Tong Tang look at another side of the coin: their area is the viral approaches to gene delivery in the setting of heart failure. They provide a summary of progress made as well as of the strengths and limitations of different viral approaches. They emphasize preclinical studies showing that gene transfer improves left ventricular contractility and attenuates deleterious remodeling in myocardial infarction–associated congestive failure, and express optimism that these outcomes will be replicated in patients with congestive failure. Finally, my own paper, authored with Peter Danilo and Richard Robinson, considers viral vector–delivered gene therapy as a means to insert novel ion channel constructs into the heart in attempting to prevent induction of lethal ventricular tachycardias. The goal here, in proof-of-concept experiments, is to provide local therapy with these constructs, thereby maintaining an antiarrhythmic action while limiting toxicity. 4 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Guest Editorial - Michael R. Rosen, MD I encourage the reader to think of the lead article and the three accompanying papers as snapshots of a field that contains a lot of empty space. We are gathering islands of information, and when we have learned a sufficient amount, the continuum of what is needed for us to understand what cell and what therapy to use in any particular situation will have been clarified. But that day is still far off. In the meantime, we learn and we worry: worry that the pressure investigators feel to bring treatments to the clinic and the economic pressures of raising funds to perform research and deliver its benefits to human subjects may poison the field by leading to premature application and unforeseen toxicity or failure. This certainly was the case for gene therapy, even though we are now having another “go” at it; it would be devastating for the same thing to happen with cell therapy. A different concern, but no less dismaying, is the administration of various cell therapies to desperately ill patients in some countries with the same abandon that characterized the selling of snake oil to a gullible public 100 years ago. Finally, I ask the reader to save this volume for about 20 years, not because of any pretensions about it, but to perform an easy experiment. The experiment is simply to open the volume in 20 years to see, with appropriate hindsight, the extent to which this bold new future we envision for gene and cell therapies has made its way into the mainstream of cardiovascular therapy. 5 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: new progress in gene/cell therapy to restore normal mechanical and electrical function Ira S. Cohen*†, MD, PhD; Glenn R. Gaudette ‡, PhD * Department of Physiology and Biophysics - Stony Brook University - NY - USA † Institute for Molecular Cardiology - Stony Brook University - NY - USA ‡Department of Biomedical Engineering - Worcester Polytechnic Institute - Worcester - Mass – USA The last decade has seen the complete sequencing of the human genome and the development of approaches to deliver therapeutic genes to the heart. Equally significant advances have also been made in stem cell biology. The accessibility of these new tools in our therapeutic arsenal raises the exciting question of whether normal mechanical and electrical function can now be regenerated in the diseased heart. In this article, we consider how to choose targets to regenerate mechanical and electrical function. In considering mechanical function, we start by describing its determinants (ie, both active and passive properties), and then review regenerative applications of gene/cell therapy. We also consider arrhythmias, focusing on the potential advantages of gene/cell therapy over pharmacotherapy or devices, and then discuss the development of biological pacemakers as one example. Overall, the future is bright; gene/cell therapy approaches have reached the proof-of-principle stage for both mechanical and electrical regeneration. Some mechanical studies have even reached clinical trials; however, evidence of long-term efficacy is lacking. Ultimately, to achieve therapeutic success with gene and cell therapies, it will be important to gain a better understanding of their mechanisms of action. n setting the stage for “Mending the Broken Heart,” it is our purpose to consider the potential of gene and/or cell therapy to restore mechanical and electrical function lost to disease. With the development of viral and stem cell technologies, new therapeutic approaches to restore normal function are possible that were not even dreamt of a decade ago. One might think that gene/cell therapy approaches would be brought to the bedside, for electrical and mechanical dysfunction, at the same rate. However, because there is no effective long-term therapy for heart I SELECTED ABBREVIATIONS AND ACRONYMS BMC bone marrow cells CHD coronary heart disease CSC cardiac stem cell CXCR4 chemokine receptor 4 Keywords: myocardial infarction; stem cell; viral vector; heart failure; biological pacemaker; myocardial regeneration Address for correspondence: Ira S. Cohen, MD, PhD, Department of Physiology & Biophysics, Health Science Center, Stony Brook University Stony Brook, NY 11790-8661, USA (e-mail: [email protected]) EB embryoid body ESC embryonic stem cell GAG glycosaminoglycan HCN hyperpolarization-activated cyclic nucleotide-gated hMSC human mesenchymal stem cell ICD implantable cardioverter-defibrillator iPSC induced pluripotent stem cell ISL1 islet 1 factor MAP mitogen-activated protein MSC mesenchymal stem cell SA sinoatrial Dialogues Cardiovasc Med. 2009;14:7-25 Copyright © 2009 LLS SAS. All rights reserved 7 www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette failure, current approaches employing gene/cell therapy to repair damage induced by myocardial infarction have moved rapidly to clinical trials, resulting in only modest success (see the article by Wollert and Drexler in this issue).1 By contrast, many existing therapies for electrical dysfunction (electronic pacemakers, implantable cardioverter-defibrillators [ICDs]) are effective and lifesaving, and, thus, the “bar” is higher, and translation to the clinic has lagged. Improvements in stent technology have allowed cardiologists to improve their treatment of myocardial ischemia. The restoration of blood flow to ischemic tissue is likely to salvage at-risk myocardium. However, for those myocytes unable to survive the insult, restoration of blood flow is of little value. The death of each myocyte decreases the potential amount of work that can be done by a region of the heart, and if a sufficient number of myocytes die, a decrease in regional work will occur. If blood flow is restored quickly, the few myocytes lost are unlikely to reduce the overall performance of the heart. For the patient who is treated too late, a significant infarct can result in decreased myocardial performance. Our aims in this article are: to briefly discuss current therapies for regenerating myocardial function; consider the targets currently under investigation; discuss the means to evaluate the success of these studies: and in the process, set the stage for new target selection. It is our purpose to emphasize “the dream” of gene/cell therapy; however, only intensive investigation in the laboratory and the clinic over the coming years will generate “the reality.” Mechanical interventions, such as balloon pumps and ventricular assist devices, currently only offer temporary solutions. The shortage of donor hearts makes heart transplantation unrealistic for many patients. The long-term solution must be the restoration of mechanical function in the heart. THE PROBLEM The World Health Organization predicts 11.1 million deaths from coronary heart disease (CHD) in the year 2020.2 In the United Kingdom, CHD is the most common cause of death, claiming 101 000 lives in 2005.2 In the same year in the United States, 16 million Americans were living with CHD, 8.1 million of whom had experienced a myocardial infarction, while 5.3 million suffered heart failure.3 In 2007 alone, about 1.2 million Americans suffered a myocardial infarction with a mortality of 451 000.3 The direct and indirect costs of heart failure in the United States are estimated at 34.8 billion dollars.3 Cardiac arrhythmias are another major affliction. In 2004, they resulted in, or contributed to, 458 800 deaths in the United States. Arrhythmias resulting from sick sinus syndrome or atrioventricular block necessitate the implantation of pacemakers, and in patients at risk of ventricular fibrillation, ICDs are indicated. There were 180 000 pacemakers and 91 000 defibrillators implanted in 2005.3 ACTIVE AND PASSIVE FUNCTION IN THE NORMAL HEART The mechanical function of the heart can be separated into active and passive components, both of which are potential targets for regenerative therapy (Figure 1). The active function results from the contraction of the myocytes, and is primarily responsible for systole. In myocardial infarction and heart failure, the myocytes are dysfunctional or destroyed; therefore, active function is decreased. However, the passive function of the ventricle also plays an important role in contraction (Figure 2). In the normal heart, cells are attached to each other and the extracellular matrix, providing a compliant environment. As the heart contracts during systole, this compliant, passive material allows for normal systolic contraction, and does not impede diastolic relaxation. If the environment is stiff, which can occur in infarcted myocardium in which myocytes and extracellular matrix are replaced by a noncompliant scar, systolic contraction is decreased. Diastolic function is also affected, as a stiff environment decreases diastolic relaxation. During contraction, the myocytes shorten, and deform the substrate to which they are anchored. If the myocytes are attached to a stiff material, such as collagenous scar tissue, it is more difficult to deform than normal extracellular matrix. This is likely to occur in nontransmural infarcts, and in the MYOCARDIAL REGENERATION While treatment for myocardial ischemia has made significant progress over the past decades, myocardial infarction remains an elusive target. Current treatments for infarction fail to address the problem at hand: the loss of cells that provide mechanical function to the heart. This portion of the review will address the functions and constraints of regenerative therapies, and evaluate their success in restoring mechanical function. 8 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette Improved mechanical function Passive material properties Cells Active material properties Proteins Exogenous Endogenous Myocyte proliferation iPS cells Embryonic stem cells Mesenchymal stem cells Cardiac stem cells border zone between infarcted and spared myocardium. Thus, one means to improve overall function is to restore the passive properties of the myocardium. wall rupture. However, scar stiffness can impede diastolic filling. Thus, increasing the compliance of the scar may lead to improved ventricular performance. It should be noted that increasing the compliance of the scar excessively has the potential to lead to aneurysm formation (Figure 3, page 10). Scar tissue is largely composed of collagen and fibroblasts. The restoration of blood flow to scar tissue may increase its compliance. For a consideration of angiogenesis employing viral therapies, see the article by Hammond and Tang in this issue.4 APPROACHES TO IMPROVING PASSIVE MECHANICAL PROPERTIES IN THE INFARCTED HEART In most cases, the body’s reaction to a myocardial infarction is scar formation. This fulfills an important need: mechanical stabilization to prevent ventricular Myocyte nt plia Com strate sub LS Myocyte contraction LD Myocyte S sub tiff stra te LS nt plia Com strate sub LD Myocyte relaxation LS Stem cell differentiation Figure 1. Means of improving mechanical function. Mechanical function in the infarcted heart can be improved through multiple approaches. However, the ultimate effect is through improved passive and/or active (contractile) properties of the infarcted region of the heart. S sub tiff stra te LD Figure 2. The passive properties of myocardium can affect deformation of the ventricular wall. In systole (top panel), a compliant material will deform along with the myocyte, leading to a systolic contraction. However, the contracting myocyte may not be able to deform the stiff substrate (for example, a stiff scar), leading to decreased systolic contraction. During diastole (bottom panel), a compliant material will expand along with the myocyte. In a stiff substrate, the expanding myocyte may not expand the substrate, leading to decreased diastolic filling. Abbreviations: LD, diastolic length; LS, systolic length. 9 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette For example, skeletal myoblasts have been shown to improve myocardial function, yet once differentiated into myotubes, they do not form gap junctions, thereby limiting their ability to contract in synchrony with spared myocardium.10 Cross-section of normal heart Stiff scar tissue Compliant scar tissue The addition of elastin to collagenous scar tissue may also improve regional mechanical function in the infarcted heart. Recent work from Li’s laboratory has demonstrated that the overexpression of elastin by endothelial cells improved cardiac function compared with nonexpressing cells.11 The addition of elastin also reduced scar size, suggesting it may have replaced the collagen, or signaled for decreased collagen synthesis. Surgically, the replacement of dysfunctional myocardial tissue can be accomplished through the endoventricular circular patch plasty procedure (also referred to as the Dor Procedure).12 While most surgeons use Dacron as patch material, the use of a more compliant scaffold may provide the opportunity to improve regional mechanical function. We have recently demonstrated that the implantation of a compliant patch (made from the isolated extracellular matrix of a porcine urinary bladder) improves regional mechanical function in contrast to a Dacron patch.13 While many other factors probably play a role in the improved function, scaffold compliance may be essential to the improved contraction observed in the patch region. Too-compliant scar tissue Figure 3. Changes in patch passive properties can lead to different filling volumes. Altering the passive properties of the ventricular wall can lead to improved global function, including increased diastolic filling. However, if the region is too compliant, ballooning similar to a ventricular aneurysm can occur. Matsubayashi et al seeded vascular smooth muscle cells onto a biopolymer scaffold to replace myocardial scar tissue in a rat model.5 Eight weeks after scaffold implantation, they noted increased extracellular elastin and increased fractional area shortening. As the patch region was akinetic, these results suggest that the improvement was not due to the restoration of contraction in the region, but probably to scaffold compliance. Implantation of a different biopolymer scaffold on the epicardial surface of a myocardial infarct by Fujimoto et al6 resulted in an increased presence of smooth muscle cells in the infarct tissue. Improved fractional area shortening was also noted in the patch group, as was compliance of the heart. Confirming the importance of passive properties, Wall et al employed a computational model of the ventricle, and demonstrated that the injection of a compliant material could, in theory, improve mechanical function.7 While increasing the passive function of infarcted myocardium may lead to improved overall heart performance, the regeneration of functional myocardium is necessary for the contraction of the heart. Regenerated myocardium must actively contract, which requires contractile cells. APPROACHES TO IMPROVING CONTRACTILE PROPERTIES IN THE INFARCTED HEART The addition of cells or compliant materials can decrease the stiffness of scarred myocardium. Berry et al recently demonstrated that the injection of mesenchymal stem cells (MSCs) after myocardial infarction leads to increased compliance.8 These cells also decreased fibrosis in the infarcted region. No evidence of differentiated MSCs was found, suggesting that nondifferentiated MSCs improve compliance. Contractile cells are needed to fully restore regional mechanical function. In myocardial infarction–associated heart failure, at least 1 billion myocytes are lost and must be replaced.14 Cells used in regenerative therapy: expected properties Interestingly, nearly any type of cell delivered to the heart appears to improve myocardial function,9 although differentiation of most cells into cardiac myocytes appears to be limited. Prior to understanding the different potential mechanisms for regenerating active function in infarcted myocardium, we need to understand the aims of a 10 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette successful regenerative strategy. Cells that are successful at regenerating functional myocardium must possess the following (adapted from reference 15): • Proteins specific to cardiac myocytes. Proteins, specific to mechanical function (eg, sarcomeric α-actinin, ventricular myosin heavy chain), electrical function (gap junctions, cardiac-specific ion channels), or electricalmechanical coupling of the myocyte, must be present. • Sarcomeres. Efficient contraction of myocytes is dependent on the organization of actin-myosin crossbridges to produce longitudinal shortening. • Ion channels to generate an action potential. Contraction is preceded by an action potential that facilitates the inflow of calcium from the extracellular space and its triggered release from the sarcoplasmic reticulum, which is essential for the contraction of the myocyte. • Functional excitation-contraction mechanisms. The generation of an action potential needs to be coupled to contraction in the myocyte. • Gap junction proteins. The ability to form gap junctions is essential for the formation of a functional syncytium, necessary for efficient activation to occur, which provides the template that orders the contraction of the heart. or allogenic cells to the heart. Embryonic stem cells (ESCs), mesenchymal stem cells, cardiac stem cells (CSCs), and induced pluripotent stem cells (iPSCs) can be expanded in culture, and then delivered to the heart. Each serves as a potential means of exogenous regeneration. Both endogenous and exogenous methods are currently under investigation. Endogenous regeneration of the heart Myocyte proliferation Common dogma suggests that cardiac myocytes lack the ability to proliferate. However, in 2001, data from Anversa’s laboratory suggested that myocytes may reenter the cell cycle in regions bordering a myocardial infarction.16 The data demonstrated that approximately 4% of the myocytes in the border zone between infarcted and viable myocardium were positive for Ki-67, a nuclear molecule involved in cell proliferation. However, after myocardial infarction, the heart does not reconstitute a sufficient number of myocytes, so the damaged tissue is replaced by scar tissue. This does not rule out the ability of myocytes to proliferate; it only suggests that myocyte proliferation is limited, and not favored over scar formation. In fact, myocyte proliferation has also been documented in other models. For example, noninfarcted zebra fish and amphibians regenerate amputated parts of the heart. This occurs as a result of mitotic expansion of cardiomyocytes.17,18 Myocyte proliferation has also been noted in a mammalian model, the medical research laboratory (MRL) mouse.19 These mutant mice regenerate wounds without forming scars, presumably due to an altered mechanism of extracellular matrix remodeling. The mitotic index of myocytes in the MRL mouse was shown to be 10% to 20% during regeneration of cryogenically injured heart.19 When delivered to the heart, these cells must also: • Increase active regional mechanical function. When the cells are delivered to an infarct, they must increase the contraction of the infarcted regions. • Regenerate myocyte mass that correlates with function. The number of regenerated cells in an infarcted region should correlate with improved mechanical function in the region, if the improved function is directly due to the cardiac differentiation of delivered cells. • Increase overall heart function. The ultimate success of regenerated myocytes is dependent on the restoration of mechanical function to the heart. Other investigators have also documented myocyte proliferation in various environments. Schuster et al induced endogenous myocyte proliferation in a rat infarct model by delivering human endothelial progenitor cells.20 By using a rat-specific antibody to Ki-67, they assured that native rat myocytes, rather than the exogenously delivered human cells, entered the cell cycle. In vitro experiments have also demonstrated the possibility of inducing myocyte proliferation. Busk et al induced myocyte proliferation through the overexpression of cyclin D2,21 a cyclin necessary for cells to progress through the G1 phase and into the S phase of the cell cycle.22 Recently, p38 mitogen–activated protein (MAP) kinase inhibition has been shown to allow adult cardiomyocytes to proliferate in vitro.23 While this list does not delineate all the functions of a successful regenerative therapy, it outlines the major accomplishments that need to be fulfilled. With these functions defined, various means can now be considered. All methods to regenerate active mechanical function must include the addition of cell mass to replace the myocytes lost to infarction. These methods can be divided into endogenous and exogenous approaches. Endogenous methods involve recruiting native cells to replenish myocyte mass. Examples would include the induction of native myocytes to proliferate, or the cardiac differentiation of native stem cells that reside in the heart, or that have homed to the infarcted tissue. Exogenous methods rely on the addition of autologous 11 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette mals, endogenous therapies have a promising future. If the appropriate environment requires native stem cells, then homing to the region of damage will be important. Multiple targets capable of optimizing homing are currently under investigation. In summary, adult mammalian cardiac myocytes can proliferate, and could, in principle, replace the myocytes lost to disease. However, to harness this potential therapy, an improved understanding of cell cycle progression in adult mammalian cardiac myocytes is necessary. Exogenous regeneration of the heart The aim of exogenous regeneration is to take a stem cell capable of becoming a myocyte in vivo, and induce it to choose a cardiac lineage. Because of the enormous number of myocytes required, and the paucity of proliferation once fully differentiated (see above section), current strategies have emphasized in vitro expansion and cardiac commitment prior to delivery. Mobilizing native stem cells The adult human possesses multipotent cells that reside in many different tissues, including bone marrow, fat, and heart. Recent evidence suggests that cells from adipose tissue,24 bone marrow (described in detail below), and heart,25 may be able to differentiate into myocytes. This property provides an opportunity to develop a therapy that will mobilize these cells, and have them home to the injured myocardium. This strategy is illustrated in Figure 4. We now consider the cell types that have been employed. Additional details on the success of cell therapy can be obtained from the article by Wollert and Drexler in this issue.1 ? Skeletal myoblasts Skeletal myoblasts, which are skeletal myocyte progenitors, were the first cellular therapy for myocardial infarction. Like cardiac myocytes, skeletal myoblasts are striated; however, their action potentials are brief in comparison with heart cells. This may account for some of the arrhythmias reported with this therapy.32 Figure 4. The goal of cell therapy in cardiac regeneration. The hope of cell therapy is to be able to deliver a cell, like the mesenchymal stem cell (left), that will become a cardiac myocyte (right). The starting cellular material and the methods needed to drive the cell down a cardiac lineage are still under investigation. While stem cells can home to the infarcted myocardium,26,27 increasing homing provides more help. Several homing molecules have been identified, including monocyte chemotactic protein-3,28 stromal-derived factor-1,29 and its receptor, chemokine receptor 4 (CXCR4).30 When skeletal myotubes differentiate from skeletal myoblasts, they do not form gap junctions.10 Without these intercellular ion channels, electrical activity cannot propagate between the endogenous cardiac myocytes and the exogenous differentiated skeletal myotubes, making it nearly impossible for the transplanted cells to contract synchronously with the native myocytes. Increased homing may also be facilitated by the addition of growth factors, which mobilize CXCR4.31 The initial data are encouraging. However, it remains a challenge to induce a sufficient number of these cells to home to an infarct, and improve mechanical function by differentiating into myocytes, or by releasing paracrine factors that induce native myocytes to proliferate, and provide appreciable recovery of mechanical function. However, delivery of skeletal myoblasts has led to improved mechanical function in animal models, probably due to improved passive function of the heart. These encouraging results have led to clinical trials.32 Some trials have shown improvements in cardiac function, although ventricular tachyarrhythmia remains a major concern. In February 2006, Genzyme halted its phase 2 clinical trials after enrolling 95 patients, citing that a significant improvement in heart function was unlikely to come from the study.33 Despite this setback, it appears that clinical trials are continuing.34 Summary of endogenous regeneration While much of the attention associated with cellular therapy for cardiac regeneration has focused on the delivery of cells to the heart, endogenous methods to regenerate heart offer additional opportunities. If the results of Keating’s laboratory, to induce myocyte proliferation in the zebra fish, can be adapted to mam- 12 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette Embryonic stem cells suggest tumor reduction may be feasible,40 clinical application will require that there be no tumor formation. Many studies have shown the ability of ESCs to differentiate into cardiomyocytes. To differentiate ESCs down a cardiac lineage, embryoid bodies (EBs) are formed. This method can include culturing cells in hanging drops of media to allow them to coalesce and form three-dimensional structures.35 After formation of the EBs, they are plated for a few weeks. Kehat et al showed that the resultant cells express cardiac proteins and approximately 8% of the EBs contract 21 days after plating.36 He et al demonstrated that some cells grown from the EBs generate action potentials, some of which resemble atrial-, ventricular- and nodal-like action potentials.37 For more on the cardiogenicity of ESCs, see the review by Laflamme and Murry.14 Bone marrow cells In 2001, Orlic et al documented regeneration of cardiac function and cardiac myocytes through the delivery of bone marrow cells (BMCs).41 These authors demonstrated the delivery of Lin– c-kit+ BMCs to the infarcted mouse heart resulted in new myocyte formation, confirmed by an enhanced green fluorescent protein tag that marked their BMCs. However, attempts by others to repeat these findings failed.42-44 Nonetheless, this initial study has spurred research into specific types of BMCs. MSCs found in the bone marrow have shown potential for cardiac differentiation. Incubation of MSCs with 5-azacytidine, a DNA-demethylation agent, results in differentiation of some of the MSCs down a cardiac lineage. The differentiated cells express cardiac specific proteins and form sarcomeres. We have recently induced MSCs to express cardiac markers by creating spheroids, a process similar to the formation of embryoid bodies.45 Approximately 15% of these cells express a calcium current similar in magnitude to that characteristic of adult ventricular myocytes. When implanted into the canine heart, some of the cells develop sarcomeres. Delivery of cardiac myocytes derived from ESCs has improved mechanical function in the injured heart. Using a myocardial infarction mouse model, Kofidis et al demonstrated that injected human ESCs improve cardiac function, although they were unable to demonstrate the presence of mature cardiomyocytes.38 Therefore, improved passive mechanical properties as a reason for improved cardiac performance cannot be ruled out. Kolossov et al were able to find ESC-derived striated cardiomyocytes after delivering ESCs to mouse heart.39 Delivery of these cells to the injured heart also significantly improved function, compared with delivery of media alone. However, to improve cell retention, it was necessary to deliver fibroblasts with the ESCderived cardiomyocytes. When comparing the fibroblast-only group with the fibroblast-plus-ESC group, it was difficult to see any difference, bringing into question the added active function benefit resulting from delivery of ESCs. Behfar et al detected sarcomere formation after delivery of cardiac-committed ESCs to the infarcted mouse heart.40 They also documented mechanical recovery of function, but did not investigate whether this was due to the contribution of the cells to active contraction or to changes in the passive properties of the tissue. In summary, evidence exists suggesting that MSCs can be induced to choose a cardiac lineage; however, generating a large quantity of a pure population of these committed cells remains a challenge. Resident cardiac stem cells Early studies from Anversa’s laboratory identified Lin– c-kit+ cells in the adult rat heart that could partially differentiate into cardiac myocytes.46 These CSCs are mobile, as they are detected in different locations throughout the infarcted heart, and they express cardiac-specific proteins. The literature is clear on the ability of ESCs to differentiate, both in vitro and in vivo, into striated cardiac myocytes. Yet data to correlate the number of engrafted ESC-derived myocytes with regional function are still lacking. In addition, complete recovery of infarcted tissue has not been shown. Other CSCs were identified, including stem cell antigen–1 (Sca-1) positive cells,47 side population cells (based on Hoechst dye exclusion),48 and islet 1 factor (ISL1).49 These different populations of stem cells appear to have little overlap (although Sca-1 positive and side population cells appear to have some overlap). All have been reported to differentiate into myocytes with contractile properties. Whether these cells exist in humans, and why they are unable to restore Aside from the political and moral debate regarding the source of ESCs, a major impediment to the use of these cells is their tumorigenesis. While recent results 13 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette normal function to the infarcted heart, is still unknown. If CSCs could be harvested and expanded, it would enhance their potential as a therapy. However, CSCs appear to be rare. In an attempt to isolate and expand CSCs, Messina et al cultured small pieces of human and mouse myocardium.25 They collected the round “phase-bright” cells that appeared to migrate from the tissue. When expanding these cells, they formed “cardiospheres,” which could be expanded. When delivered to the infarcted mouse heart, cells from cardiospheres improved ejection fraction compared to the delivery of fibroblasts.50 the heart. Current routes for delivering cells used for regenerative therapy are intravascular, intracoronary, and intramyocardial. While intravascular delivery of cells is the least invasive, most of the cells are trapped in the lungs,27 with less than 1% of the cells residing in the infarcted heart.54 During angioplasty, cells can be delivered intracoronarily, directly to the region of interest. However, upon restoration of blood flow, the majority of cells are washed away from the region of interest, and only 3% of those delivered are engrafted into the heart.55 The intramyocardial route for injection of cells resulted in 11% engrafting in the heart.55 As with other cell types, the percentage of CSCs that differentiate down a cardiac lineage is unknown, as is the contribution of the differentiated cells to improvements in mechanical function. Hence, the possibility of isolating a clinically applicable quantity of pure cardiac myocyte precursor cells with this approach is still questionable. While many researchers have developed tissue constructs that incorporate fetal or neonatal rat cardiac myocytes into engineered cardiac tissue (see Radisic et al, 2007 for review56), only a few of them have investigated scaffold-based strategies for delivering stem cells to the heart. Materials used include alginate,57 collagen,58 collagen/glycosaminoglycan (GAG),59 and Matrigel.60,61 However, stem cells delivered via scaffolds appear to have a difficult time traversing the myocardial wall to reach the endocardium,58 where many clinical myocardial infarctions reside. Recently, Simpson et al, using a scaffold-based delivery vehicle, demonstrated that only 1% of engrafted human mesenchymal stem cells (hMSCs) reside in the endocardial space. Induced pluripotent stem cells Current beliefs suggest that once a cell has differentiated, it cannot completely turn back. However, recent evidence from Takahashi et al demonstrated that human cells can be reprogrammed to acquire ESC-like properties.51 In summary, using current methods, it is difficult to efficiently deliver a large number of stem cells to a well-defined region where they will be retained. In the absence of an improvement in cell retention, it will be difficult for any exogenous regenerative approach to succeed. By transfecting human dermal fibroblasts with Oct3/4, Klf4, Sox2, and c-Myc, they were able to induce the expression of several ESC-specific markers, suggesting the derivation of iPSCs. Two groups were recently able to derive cardiac cells from iPSCs. Narazaki et al induced vascular cell markers from iPSCs.52 In addition, by coculturing iPSCs with OP9 stromal cells, they could induce cardiac-specific markers and striated cells. In addition, some of these cells generated a nodal-like action potential. Using an EB-like differentiation process, Mauritz et al generated contracting EBs.53 Some of these cells also expressed cardiac-specific markers and were striated. These cells also expressed the gap junction protein, connexin 43, and exhibited functional coupling. Major concerns that need to be addressed with iPSCs are the gene infection required to induce pluripotency and the potential for tumorigenesis. Summary of exogenous regeneration The search for a pure population of cells that can differentiate into myocytes continues. While embryonic stem cells appear to differentiate reliably into myocytes, issues with tumorigenesis continue to be a major concern. Other cell types have been shown to differentiate into myocytes, but none on a consistent basis in large enough quantities to effectively restore myocardial function. Indeed, there is no reported correlation between the number of new myocytes and the change in regional mechanical function, a necessary criterion to evaluate success. In addition, efficient delivery of any cell to the heart remains elusive. Delivery of exogenous cells Another issue complicating exogenous methods of cardiac regeneration is the delivery of these cells to 14 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette The pacemaker potential is a slow depolarization generated by a net inward current flow. In the normal heart, the action potential propagates from the primary pacemaker cells in the SA node, to the most distal cells in the ventricular epicardium by current flow through intercellular channels, called gap junctions.62 While gene therapy can directly deliver genetic material to myocytes, cell therapy requires electrical integration of the delivered cells. This is most often accomplished by gap junction formation between delivery and target cells. RESTORING MECHANICAL FUNCTION TO THE INFARCTED HEART Restoring mechanical function to the heart appears to be achievable, at least through passive mechanisms. However, to fully regenerate myocardium, a significant quantity of contractile cells must be produced. Neither endogenous nor exogenous methods have been able to fully restore mechanical function to the infarcted heart. Although hopeful signs for regeneration exist, it remains to be seen whether myocyte proliferation or stem cell differentiation will make functional myocardial regeneration a clinical reality. COMPARISON OF PHARMACOTHERAPY AND DEVICE THERAPY WITH GENE/CELL THERAPY ELECTRICAL REGENERATION WITH GENE/CELL THERAPY The last half-century of pharmacological discovery has focused on development of small molecules capable of influencing excitability. Device therapy has focused on the development and implantation of electronic pacemakers and cardioverter-defibrillators, as well as on surgical or catheter ablation of arrhythmogenic foci, or interruption of reentrant pathways. A comparison of gene/cell therapy with either pharmacotherapy or device therapy suggests some potential advantages of the former: The heart is a rhythmic pump, whose mechanical function requires orderly electrical activation. In this section, we lay out the reasons why gene/cell therapy has been considered as an alternative, and describe the means by which targets are selected. It should be noted at the outset that there are fundamental differences in approaches to gene/cell therapies for mechanical and for electrical regeneration. When the heart fails as a rhythmic pump, it is a global problem, and the optimal solution requires the replacement of a billion or more myocytes. Arrhythmias, in contrast, can often be treated focally by influencing a million or fewer cells. Pharmacotherapy • Specificity. Most pharmacologic agents are nonspecific. In addition to their actions on the channel/transporter of interest, they frequently have actions on others. For example, Class I drugs that slow or terminate conduction via Na channel blockade can also block K channels. Class III drugs that prolong refractory period do so by prolonging repolarization as well. Excess prolongation of repolarization can be proarrhythmic. • Types of actions. There are far more channel or transporter blockers than there are openers, however, more often than not, the electrical abnormalities that generate arrhythmias are due to reduced ion channel/transporter activity. For example, in the border zone of an infarct the myocytes are depolarized, inactivating Na channels and reducing Na conductance on depolarization, resulting in slow conduction that predisposes to reentry. • Targets. Drugs are limited to actions on those channels or transporters already present in the myocytes. That is, one cannot “import” a target into the cell that then is subject to therapeutic drug action. NORMAL ELECTRICAL ACTIVITY: THE ACTION POTENTIAL AND PACEMAKER ACTIVITY The electrical activity of the heart, delivered via orderly propagation of the cardiac action potential, precedes and initiates the mechanical event, the contraction. All the ion channels and transporters necessary to initiate the heartbeat reside in the membranes of the sinoatrial (SA) node myocytes. The nervous system modulates this activity, but does not initiate it.62 The action potential is generated by a population of ion channels and transporters, each of which is comprised of one or more proteins generating either inward or outward current across the myocyte membrane. The voltage waveform of the action potential is the result of the summed ion movements through all the channels and transporters of a cell. A net inward current (positive ions moving into the cell) generates a membrane depolarization, while a net outward current (positive ions moving out of the cell) generates a membrane repolarization. The sum total of these limitations means that pharmacologic therapy for electrical dysfunction is not usually aimed at restoring normal function, but at limiting the 15 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette consequences of abnormal electrical activity instead, by making offending regions electrically unexcitable. • Origin of targets. The entire human genome has been sequenced. Therefore, channels/transporters expressed in the heart can be used to correct abnormalities, as can those normally located in other tissues. Furthermore, if the properties of native channels are less than optimal, then a more optimal gene can be created by modifying existing genes or by creating new ones by genetic engineering. Device therapy • Maintenance. Pacemakers require regular maintenance due to battery rundown and occasional lead fracture. • Autonomic responsiveness. Pacemakers are not autonomically responsive. However, rate-responsive units do increase rate in response to the demands of exercise. • Placement. It would be ideal to position electrodes at sites that maximize mechanical performance. Yet, this cannot be routinely done. Even using biventricular pacing, electrode positioning is limited by the anatomy of the coronary venous system. • Inappropriate responses. Inappropriately delivered shocks are a complication of ICDs that can cause physical and psychological trauma. • Venous stricture and recurrence of arrhythmia. Atrial fibrillation originating from the pulmonary veins is often treated with ablation. A complication of this therapy is venous stricture. Furthermore, antiarrhythmic success with ablation can be complicated by later recurrence. When compared with device therapy: • Maintenance. No maintenance is required. • Autonomic responsiveness. Normal sympathetic and parasympathetic responsiveness should occur as long as innervation is present at the site of implantation. • Placement. With regard to biological pacemakers, positioning of the implant by catheter delivery can be used to optimize mechanical performance of the ventricles. • Inappropriate responses. These can be terminated either by regulated expression of the exogenous gene (the promoter-inducing expression can be turned off), by pharmacotherapy that blocks the overexpressed channel/transporter protein, or by ablation of the site. There is minimal tissue damage with delivery of either gene or cell therapy. • Venous stricture and recurrence of arrhythmia. There is minimal tissue damage with delivery of either gene or cell therapy. To sum up, pacemakers were the panacea of 20th-century therapy for some disorders of rate and rhythm. However, maintenance is required, and rate responsiveness is an imperfect solution to the absence of autonomic control. ICD implantation has saved tens of thousands of lives, but patients are exposed to both physical and psychological trauma from inappropriate firing. Ablation therapy has worked for arrhythmias where other alternatives have failed, however, it carries risks of anatomical damage, and is not always effective in the long-term. In summary, gene/cell therapy has the capacity to be more specific than pharmacotherapy, to enhance as well as reduce channel function, and to employ a larger array of targets optimized by genetic engineering. Compared with devices, gene/cell therapy is maintenance-free, autonomically responsive, and optimized for mechanical performance. Inappropriate responses can be prevented or reversed by regulating exogenous gene expression, or with appropriate pharmacotherapy, and no anatomical damage should occur with deployment. In short, it has the capacity to restore normal electrical function. Gene/cell therapy We will now consider whether gene/cell therapy offers the possibility of achieving a more successful therapy. When compared with pharmacotherapy: • Specificity. A gene encodes for a single protein and, as such, is specific. There can be nonspecific effects associated with overexpression or silencing of a specific protein, but, ideally, the only action of the delivered genetic material is to alter expression of a single protein. • Types of actions. Channel/transporter activity can be enhanced by successful delivery of its cDNA, or reduced by a dominant negative construct, antisense, or small interfering RNA. CHOOSING A DELIVERY SYSTEM There are three approaches to delivering a therapeutic electrical change to the heart: • The first is conceptually the simplest. The cDNA for the channel/transporter is delivered in a naked plasmid. However, the transfection efficiency is low and the action very transient. 16 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette • Second, the gene of interest can be packaged in a virus. The delivery can be either localized 63 or to the chamber as a whole,64 a decision dictated by both the safety and the efficacy of each approach. A second decision is which virus to choose. Adenoviruses are largely safe and induce high levels of expression of the transgene, but are not persistent because they are episomal (not incorporated into the genome). Lentiviruses are more persistent because of genome incorporation, but have higher risks of neoplasia. The choice of which particular virus, as well as the properties of adeno-associated viruses, are discussed in detail in the article by Hammond and Tang4 in this issue. • The third is cellular delivery. This approach is, by definition, focal, and requires that the chosen delivery cell be able to electrically influence the target cell. In most cases, this is achieved through the formation of gap junctions. These are formed from subunit proteins, called connexins. HMSCs and ESC-derived cardiac myocytes make connexins, and can form functional gap junctions with cardiac myocytes as well as with each other, in vitro and in vivo.65-68 GENERAL PRINCIPLES OF BIOLOGICAL PACEMAKER DESIGN Figure 5 (page 18) shows the action potentials of a pacemaker cell from the SA node, and that of a ventricular myocyte. In the sinus node, a small net inward current during diastole generates the spontaneous pacemaker depolarization, while in the ventricle and much of the atrium, there is no net current flow (and thus no change in membrane potential) between action potentials. Figure 5 also indicates the major membrane conductances responsible for the pacemaker activity in the SA node myocyte, and for quiescence in the ventricular cell. If is an inward current (carried largely by Na+), activated by hyperpolarization in the sinus node. This “pacemaker” current is absent in the physiologic voltage range in ventricular myocytes. IK1 is generated by a large outward conductance present in ventricular myocytes, but is largely absent in the pacing cells of the SA node.72,73 Whether placed in the atrium or in the ventricle, the challenge in creating a biological pacemaker is to convert a diastolic interval with zero net current flow into a period of small net inward current. Finally, fusing delivery and target cells is possible. Cho et al69 transfected lung fibroblasts and induced fusion of these fibroblasts to myocytes in the guinea pig heart. This approach avoids the potential loss of function during ischemia where gap junctions might tend to uncouple. However, the long-term effects of cell fusion as well as the agent that induces it (polyethylene glycol) remain unknown. Current gene and cell therapy approaches to this problem are illustrated schematically in Figure 6 (page 19) and discussed in detail below. Reducing background outward In their article in this issue, Rosen et al70 discuss approaches of gene/cell therapy to life-threatening tachyarrhythmias. It is clear that native cardiac, mutated cardiac, and even noncardiac genes are showing proof of principle as therapies in animal studies in that arena. Below, we discuss an alternative to devices: biological pacemakers. IK1 current leads to pacemaker activity in normally quiescent myocytes Since IK1 is the dominant conductance during diastole in ventricular myocytes, reducing this outward current in normally quiescent tissue should result in a net inward current generating a pacemaker-like membrane depolarization toward threshold. Miake et al64 used a dominant negative form of Kir2.1 (a molecular correlate of the IK1 channel), and an adenoviral delivery system in the guinea pig ventricle, to reduce the number of functioning IK1 channels. STRATEGIES FOR THE DEVELOPMENT OF BIOLOGICAL PACEMAKERS The area showing the widest array of conceptual approaches to gene/cell therapy has been the development of biological pacemakers. Beginning in 1998,71 a number of investigators have used a variety of approaches to create biological pacemakers as an alternative to these electronic devices. These approaches display not only some of the principles of gene/cell therapy design, but also point out some of the limitations of this approach. This approach works in the manner illustrated in Figure 7 (page 19). The IK1 channels, like most K channels, are composed of 4 identical subunits. A dominant negative construct is a genetically modified form of the channel subunit, which combines with the native functional channel subunits to form nonfunctional channels that do not permit current flow. The disadvantage of such an approach is illustrated in Figure 5 17 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette Sinus node myocyte Net inward current If 1 us I K Min Ventricular myocyte Plu sI f Zero net current IK1 Figure 5. Pacing requires net inward current. Schematic action potentials from a sinus node (top left panel) and a ventricular myocyte (bottom left panel). The sinus node myocyte paces, and, thus, has a net inward current flowing during diastole, while the ventricular myocyte reaches a constant “resting potential” because the sum of all currents flowing across the membrane is zero. The major “pacemaker” current in the sinus node is If , while the dominant conductance setting the resting potential in the ventricular myocyte generates a background outward current, called IK1. Panels on the right show approaches to creating a biological pacemaker by removing IK1 from, or adding If to, the ventricular myocyte. IK1 flows during the final repolarization as well as during diastole, so decreasing its magnitude lengthens the action potential (top right panel: control action potential in orange, minus IK1 in green). Since If is activated only during diastole, adding it has little or no influence on the action potential duration (lower right panel: control action potential in orange, plus If in green). Both approaches create a biological pacemaker, but removing IK1 results in a longer QT interval. (upper right panel). IK1 contributes to final repolarization of the ventricular action potential. Thus, in addition to inducing pacemaker activity, delivery of this dominant negative form of IK1 also prolongs the action potential and the QT interval.74 maker potential in the sinus node is called If,73 and is not activated at diastolic potentials in quiescent atrium or ventricle. The α-subunit of this channel is encoded by the hyperpolarization-activated–cyclic nucleotidegated (HCN) gene family.75,76 As its name indicates, this channel opens when the action potential repolarizes to diastolic membrane potentials. Since it closes rapidly on depolarization,77 its action is limited to the diastolic time period, avoiding the problems experienced with downregulation of IK1 (discussed above). An important lesson learned from this approach is that individual channel types can have effects on different phases of the action potential. In this case, the reduction of IK1 had the desired effect of producing net inward current during diastole. However, it also had the undesired effect of increasing action potential duration. Another advantage of this approach is that the portion of the If channel in the cytoplasm binds the cyclic nucleotide cAMP,78,79 leading to the opening of more If channels upon sympathetic stimulation.80 This creates the autonomic responsiveness desired in a “biological pacemaker.” In our initial studies, the HCN2 gene (one member of this multigene family) was delivered to the canine left atrium81 or left bundle branch63 by adenovirus, or to the left ventricular free wall in transfected human MSCs,65 where it either created (in the atrium or ventricle), or enhanced (left bundle branch), native pacemaker function. Persistent biological pacemaker function was demonstrated for 6 weeks when at least Adding inward pacemaker current also creates pacemaker activity Native HCN genes If net inward current is required for pacemaker activity, the obvious alternative approach to reducing an outward current is to add an inward current (Figure 5, lower right panel). As stated above, the pacemaker channel that carries the major inward current during the pace- 18 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette A Increase net inward current during diastole β2-Adrenergic receptors Engineered HCN genes B Decrease outward IK1 Increase inward If Increase sympathetic tone Genetically modified Kv1.4 channel Native HCN genes Dominant negative Kir2.1 Increase net inward current during diastole Autologous SA node myocytes hMSCs transfected with HCN genes ESCs differentiated to pacemaker myocytes Fibroblasts transfected with an HCN gene Integration via gap junctions Integration via gap junctions Integration via gap junctions Integration via cell fusion Figure 6. Gene and gene/cell therapy approaches to creating a biological pacemaker. A. Gene therapy approaches. B. Approaches that use either cell therapy or a combination of gene and cell therapy. Details provided in text. Abbreviations: HCN, hyperpolarizationactivated–cyclic nucleotide-gated. 700 000 hMSCs (roughly half carrying the HCN2 gene) were delivered to the canine left ventricular free wall.82 The hMSCs carrying the gene showed no evidence of humoral or cellular rejection, and the rhythm was catecholamine sensitive. The absence of hMSC rejection in this xenograft was not a complete surprise, as these cells are known to possess some immune privilege.82 Although native genes demonstrated both acceptable basal rates (roughly 60 beats per minute) and some increase in rate, in response to catecholamines, neither was optimal. Modified HCN genes If the native pacemaker genes are not optimal, it is possible an improved channel form could be created by genetic engineering. When a new ion channel is cloned, investigators study the relationship of its structure to its function.80 Such was the case for the HCN gene family, in which the portion of the channel responsible for its voltage dependence, its kinetics, and its cAMP binding were defined.80,83 To increase the rate of pacemaker activity, more If channels must open during diastole. This can be achieved by shifting the voltage dependence of opening to more positive po- Figure 7. Dominant negative suppression of ion channel function. Upper panel: A representation of a longitudinal section through a functional channel (top right) in a membrane (green), composed of 4 endogenous subunits (orange) that allow ions (violet) to pass through the pore. A nonfunctional channel (top left) comprised of 3 endogenous subunits (orange) and 1 exogenous (dominant negative) subunit (grey). The grey barrier is there to illustrate that the nonfunctional channel does not allow ions (violet) to permeate. Lower panel: Cross-section through a functional (4 orange subunits), and a nonfunctional (3 orange subunits and 1 grey subunit) channel. These illustrate that 4 subunits come together to form the channel and create a pore. 19 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette tentials, or by speeding the kinetics of channel opening. We chose a mutant HCN channel with a more positive voltage dependence, but it expressed more poorly in myocytes than the native channel.84 There was no increase in basal pacemaker rate, although there was enhanced catecholamine sensitivity. Tse et al85 used a mutated form of the most rapidly activating HCN isoform, HCN1, whose voltage dependence favored channel opening. They delivered it by adenovirus to the pig atrium (where the opposing IK1 current is smaller than in ventricle). They were able to achieve physiologic rates in that location, with some autonomic responsiveness. If adding more If speeds pacemaker rate, is there any limit to this approach? The theoretical answer is yes. Since If is an inward current, it could cause a steady depolarization making cells unexcitable. The same group produced larger magnitudes of If with the same HCN1 mutant, and demonstrated that too much pacemaker current can lead to termination of pacemaker function.86 potentials, as a template. They mutated the channel to change its selectivity, converting it from K-selective to nonselective. This resulted in a change in direction of the current flowing through the channel, from net outward to net inward. The native channel opened on depolarization. They engineered the channel to open on hyperpolarization. This synthetic “HCN-mimic” was delivered by adenovirus to the guinea pig ventricle where it generated a biological pacemaker. This tour de force of genetic engineering demonstrated that existing channels could be radically reengineered to produce pacemaker function. Unfortunately, unlike the native HCN channels, this synthetic channel does not possess a cAMP binding site, and so is unlikely to be directly regulated by the autonomic nervous system. However, autonomic regulation might be achieved through effects on other ion channels/transporters present in the infected myocytes. As stated above, the other approach to increase If is to accelerate the kinetics of channel opening. We employed a chimeric channel formed from HCN1 and HCN2.87 The piece from HCN1 guaranteed faster kinetics than HCN2, while the portion from HCN2 guaranteed better cAMP-sensitivity than HCN1.83 This chimeric HCN1/HCN2 channel was delivered by adenovirus to the canine left bundle branch where it generated a ventricular tachycardia. Fortunately, this arrhythmia was eliminated by the If -blocker ivabradine, demonstrating the feasibility of terminating runaway biological pacemaker activity.87 Overexpression of the 2-adrenergic receptor Alternative gene/cell therapy approaches The first biological pacemaker was created by delivery of the cDNA for the β2-adrenergic receptor to the murine right atrium.71 Expression of this receptor increased pacemaker rate by 40%. The disadvantage of this approach was that the overexpression of a single protein had a nonspecific action. This was because a myriad of ion channels/transporters, as well as other cellular functions, are regulated by cAMP. This approach demonstrated proof of principle for biological pacing, but was potentially arrhythmogenic, so unlikely to progress to a therapeutic modality. Embryonic stem cells Thus, the lesson learned from these approaches is that more inward current is not always better. Even if the action of a gene/cell therapy is limited to the diastolic range of potentials, the amount of inward current can overshoot the optimum, leading to either arrhythmias or quiescence. As discussed in the section on mechanical regeneration, ESCs can be forced into a cardiac lineage by forming three-dimensional structures, called embryoid bodies (EBs). Cells within these cardiogenic EBs are spontaneously active. Kehat et al67 transplanted 40 to 150 spontaneously beating EBs into the posterolateral wall of the pig left ventricle to create a biological pacemaker. In those animals in which persistent pacemaker function was demonstrated, the rate was similar to the idioventricular rate of native secondary pacemakers (roughly 60 bpm). It was also responsive to adrenergic stimulation. The inward current added must be titrated against the outward current present in the delivery location to achieve the optimal pacemaker rate. Synthetic genes carrying inward current When using HCN genes in a cardiac region that natively expresses them, channels formed of native and overexpressed subunits may have unpredictable properties.88 To avoid this potential difficulty, Kashiwakura et al89 used a voltage-dependent K channel (Kv1.4) that would generate net outward current at diastolic The major advantage of this approach is that all the elements necessary for generating the pacemaker potential are included in the delivered cell. 20 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette However, the disadvantages include the risk of neoplasia from undifferentiated embryonic stem cells, and the need for immunosuppression to prevent the rejection of those embryonic stem cells that had differentiated. LOOKING TO THE FUTURE The title of our article raises the question, “Can gene/ cell therapy restore normal mechanical and electrical function”? The answer is not a simple yes or no. Proof of principle has been demonstrated for a wide variety of approaches, in both the mechanical and electrical arena, and this has raised hopes. However, definitive therapies remain an elusive target. Part of the problem is theoretical, as clear objective criteria for success have not been defined. The other part is technical, and relates to the scale of the mechanical problem requiring billions of myocytes, and the complex nature of the electrical problem due to the interactions of multiple time- and voltage-dependent ion currents. Progress can be accelerated if studies reporting measures of success also pursue an understanding of the mechanism of action. However, given the new tools in our therapeutic arsenal, the future looks bright. We have come a long way in a short time. Myriad targets show promise, and many therapies may be just beyond the horizon. How long we take to reach that point will be determined by the rigor of our investigations and the support provided by our funding institutions. Autologous SA node myocytes If the aim of biological pacemakers is to simulate sinus node function, it is natural to ask whether autologous transplantation of sinus node myocytes into a ventricular location could generate a functional biological pacemaker. Zhang et al90 successfully removed and dissociated myocytes from the canine SA node, and then delivered roughly 500 000 of these cells to the right ventricular subepicardial free wall of the same animals. Three weeks after implantation, an idioventricular rhythm of approximately 60 beats per minute was observed. The rate increased on catecholamine infusion. However, neither the basal rate nor the catecholamine responsiveness was better than that observed with HCN2-transfected hMSCs delivered to the canine left ventricular free wall, or with adenoviral delivery of HCN2 to the canine left bundle branch.63,82 CONCLUSIONS FOR ELECTRICAL REGENERATION The authors were supported by NIH grants HL28958 and HL67101, an Institutional grant from NYSTEM (ISC), and a National Scientist Development Grant from the American Heart Association (GRG). The authors would also like to thank Drs Howard L. Cohen, Richard T. Mathias, and Chris Clausen for helpful discussions. The 20th century brought us lifesaving pharmacologic and device therapies for disorders of rate and rhythm. However, these therapies are not perfect. Drugs are nonselective and, because of this, can be proarrhythmic. Devices also come with a price: regular maintenance, inappropriate responses, and anatomic damage. Gene/cell therapy has the potential to be more selective, and has a wider array of targets, than pharmacologic agents. It also has the potential to be maintenance-free and more biologically responsive than devices, yet without their associated anatomical damage. As stated in the introduction, our aim was to show you the “dream” of gene/cell therapy. Our current “reality” is nowhere near this ideal. Although “proof of principle” has been demonstrated for some gene/cell therapies for both tachyarrhythmias and bradyarrhythmias, these therapies are far from perfect. The electrical activity of the heart is dependent on the complex interactions of many ion currents. Unfortunately, because of these interactions, influencing the expression of even a single channel type can often result in outcomes that influence the activity of others, resulting in proarrhythmia. Although the successes of gene/cell therapy for electrical dysfunction are modest to date, they hold the hope of a return to normal function that must be pursued. REFERENCES 1. Wollert K, Drexler H. Does cell therapy for myocardial infarction and heart failure work? Dialogues Cardiovasc Med. 2008;14:35-41. 2. International Cardiovascular Disease Statistics. In: American Heart Association. www.americanheart.org/presenter.jhtml?identifier=3001008. Accessed December 30, 2008. 3. Rosamond W, Flegal K, Furie K, et al. Heart disease and stroke statistics—2008 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation. 2008;117:e25-e146. 4. Hammond HK, Tang T. Gene therapy for myocardial infarcted-associated congestive heart failure: how far have we got? Dialogues Cardiovasc Med. 2008;14:27-34. 21 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette 5. Matsubayashi K, Fedak PW, Mickle DA, Weisel RD, Ozawa T, Li RK. 16. Beltrami AP, Urbanek K, Kajstura J, et al. Evidence that human cardiac myocytes divide after myocardial infarction. Improved left ventricular aneurysm repair with bioengineered vascular smooth muscle grafts. N Engl J Med. 2001;344:1750-1757. Circulation. 2003;108(suppl 1):II219-II225. 17. Flink IL. 6. Fujimoto KL, Tobita K, Merryman WD, et al. An elastic, biodegradable cardiac patch induces contractile smooth muscle and improves cardiac remodeling and function in subacute myocardial infarction. Cell cycle reentry of ventricular and atrial cardiomyocytes and cells within the epicardium following amputation of the ventricular apex in the axolotl, Amblystoma mexicanum: confocal microscopic immunofluorescent image analysis of bromodeoxyuridine-labeled nuclei. J Am Coll Cardiol. 2007;49:2292-2300. Anat Embryol (Berl). 2002;205:235-244. 7. Wall ST, Walker JC, Healy KE, Ratcliffe MB, Guccione JM. 18. Poss KD, Wilson LG, Keating MT. Theoretical impact of the injection of material into the myocardium: a finite element model simulation. Science. 2002;298:2188-2190. Heart regeneration in zebrafish. Circulation. 2006;114:2627-2635. 19. Leferovich JM, Bedelbaeva K, Samulewicz S, et al. Heart regeneration in adult MRL mice. 8. Berry MF, Engler AJ, Woo YJ, et al. Proc Natl Acad Sci U S A. 2001;98:9830-9835. Mesenchymal stem cell injection after myocardial infarction improves myocardial compliance. 20. Schuster MD, Kocher AA, Seki T, et al. Am J Physiol Heart Circ Physiol. 2006;290:H2196-H203. 9. Murry CE, Reinecke H, Pabon LM. Myocardial neovascularization by bone marrow angioblasts results in cardiomyocyte regeneration. Regeneration gaps: observations on stem cells and cardiac repair. Am J Physiol Heart Circ Physiol. 2004;287:H525-H532. J Am Coll Cardiol. 2006;47:1777-1785. 21. Busk PK, Hinrichsen R, Bartkova J, et al. Cyclin D2 induces proliferation of cardiac myocytes and represses hypertrophy. 10. Mills WR, Mal N, Kiedrowski MJ, et al. Stem cell therapy enhances electrical viability in myocardial infarction. Exp Cell Res. 2005;304:149-161. J Mol Cell Cardiol. 2007;42:304-314. 22. Regula KM, Rzeszutek MJ, Baetz D, Seneviratne C, Kirshenbaum LA. 11. Mizuno T, Yau TM, Weisel RD, Kiani CG, Li RK. Elastin stabilizes an infarct and preserves ventricular function. Therapeutic opportunities for cell cycle re-entry and cardiac regeneration. Circulation. 2005;112:I81-I88. Cardiovasc Res. 2004;64:395-401. 12. Athanasuleas CL, Stanley AW Jr, Buckberg GD, Dor V, DiDonato M, Blackstone EH; RESTORE Group (Reconstructive Endoventricular Surgery, Returning Torsion Original Radius Elliptical Shape to the LV). 23. Engel FB, Schebesta M, Duong MT, et al. p38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes. Surgical anterior ventricular endocardial restoration (SAVER) in the dilated remodeled ventricle after anterior myocardial infarction. Genes Dev. 2005;19:1175-1187. J Am Coll Cardiol. 2001;37:1199-1209. 24. Planat-Benard V, Menard C, Andre M, et al. 13. Kochupura PV, Azeloglu EU, Kelly DJ, et al. Spontaneous cardiomyocyte differentiation from adipose tissue stroma cells. Tissue-engineered myocardial patch derived from extracellular matrix provides regional mechanical function. Circ Res. 2004;94:223-229. Circulation. 2005;112:I144-I149. 25. Messina E, De Angelis L, Frati G, et al. 14. Laflamme MA, Murry CE. Regenerating the heart. Isolation and expansion of adult cardiac stem cells from human and murine heart. Nat Biotechnol. 2005;23:845-856. Circ Res. 2004;95:911-921. 15. Schuldt AJ, Rosen MR, Gaudette GR, Cohen IS. 26. Jiang WH, Ma AQ, Zhang YM, et al. Repairing damaged myocardium: evaluating cells used for cardiac regeneration. Migration of intravenously grafted mesenchymal stem cells to injured heart in rats. Curr Treat Options Cardiovasc Med. 2008;10:59-72. Sheng Li Xue Bao. 2005;57:566-572. 22 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette 27. Kraitchman DL, Tatsumi M, Gilson WD, et al. 38. Kofidis T, Lebl DR, Martinez EC, Hoyt G, Tanaka M, Robbins RC. Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction. Circulation. 2005;112:1451-1461. Novel injectable bioartificial tissue facilitates targeted, less invasive, large-scale tissue restoration on the beating heart after myocardial injury. 28. Schenk S, Mal N, Finan A, et al. Circulation. 2005;112:I173-I177. Monocyte chemotactic protein-3 is a myocardial mesenchymal stem cell homing factor. 39. Kolossov E, Bostani T, Roell W, et al. Stem Cells. 2007;25:245-251. Engraftment of engineered ES cell-derived cardiomyocytes but not BM cells restores contractile function to the infarcted myocardium. 29. Aghi M, Cohen KS, Klein RJ, Scadden DT, Chiocca EA. J Exp Med. 2006;203:2315-2327. Tumor stromal-derived factor-1 recruits vascular progenitors to mitotic neovasculature, where microenvironment influences their differentiated phenotypes. 40. Behfar A, Perez-Terzic C, Faustino RS, et al. Cancer Res. 2006;66:9054-9064. J Exp Med. 2007;204:405-420. 30. Robertson P, Means TK, Luster AD, Scadden DT. 41. Orlic D, Kajstura J, Chimenti S, Bodine DM, Leri A, Anversa P. Cardiopoietic programming of embryonic stem cells for tumor-free heart repair. CXCR4 and CCR5 mediate homing of primitive bone marrow-derived hematopoietic cells to the postnatal thymus. Transplanted adult bone marrow cells repair myocardial infarcts in mice. Exp Hematol. 2006;34:308-319. Ann N Y Acad Sci. 2001;938:221-229. Discussion 229-230. 31. Mahmud N, Patel H, Hoffman R. 42. Nygren JM, Jovinge S, Breitbach M, et al. Growth factors mobilize CXCR4 low/negative primitive hematopoietic stem/progenitor cells from the bone marrow of nonhuman primates. Bone marrow-derived hematopoietic cells generate cardiomyocytes at a low frequency through cell fusion, but not transdifferentiation. Nat Med. 2004;10:494-501. Biol Blood Marrow Transplant. 2004;10:681-690. 43. Murry CE, Soonpaa MH, Reinecke H, et al. 32. Menasche P, Hagege AA, Vilquin JT, et al. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Autologous skeletal myoblast transplantation for severe postinfarction left ventricular dysfunction. Nature. 2004;428:664-668. J Am Coll Cardiol. 2003;41:1078-1083. 44. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. 33. Genzyme. Genzyme Ends Enrollment in Phase 2 Trial of Cardiac Cell Therapy. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. 2006. Nature. 2004;428:668-673. 34. Menasche P. 45. Potapova I, Doronin S, Kelly D, et al. Current status and future prospects for cell transplantation to prevent congestive heart failure. Enhanced recovery of mechanical function in the canine heart by seeding an extracellular matrix patch with mesenchymal stem cells committed to a cardiac lineage. Semin Thorac Cardiovasc Surg. 2008;20:131-137. Am J Physiol Heart Circ Physiol. 2008;295:H2257-H2263. 35. Boheler KR, Czyz J, Tweedie D, Yang HT, Anisimov SV, Wobus AM. 46. Beltrami AP, Barlucchi L, Torella D, et al. Differentiation of pluripotent embryonic stem cells into cardiomyocytes. Adult cardiac stem cells are multipotent and support myocardial regeneration. Circ Res. 2002;91:189-201. Cell. 2003;114:763-776. 36. Kehat I, Kenyagin-Karsenti D, Snir M, et al. 47. Oh H, Chi X, Bradfute SB, et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. Cardiac muscle plasticity in adult and embryo by heart-derived progenitor cells. J Clin Invest. 2001;108:407-414. Ann N Y Acad Sci. 2004;1015:182-189. 37. He JQ, Ma Y, Lee Y, Thomson JA, Kamp TJ. 48. Pfister O, Mouquet F, Jain M, et al. Human embryonic stem cells develop into multiple types of cardiac myocytes: action potential characterization. CD31- but not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Circ Res. 2003;93:32-39. Circ Res. 2005;97:52-61. 23 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette 49. Laugwitz KL, Moretti A, Lam J, et al. 61. Laflamme MA, Chen KY, Naumova AV, et al. Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nature. 2005;433:647-653. Nat Biotechnol. 2007;25:1015-1024. 50. Smith RR, Barile L, Cho HC, et al. 62. Noble D. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Initiation of the Heart Beat. 2nd edition. Oxford, UK: Clarendon Press; 1979. Circulation. 2007;115:896-908. 63. Plotnikov AN, Sosunov EA, Qu J, et al. 51. Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Biological pacemaker implanted in canine left bundle branch provides ventricular escape rhythms that have physiologically acceptable rates. Cell. 2007;131:861-872. Circulation. 2004;109:506-512. 52. Narazaki G, Uosaki H, Teranishi M, et al. 64. Miake J, Marban E, Nuss HB. Directed and systematic differentiation of cardiovascular cells from mouse induced pluripotent stem cells. Biological pacemaker created by gene transfer. Nature. 2002;419:132-133. Circulation. 2008;118:498-506. 65. Potapova I, Plotnikov A, Lu Z, et al. 53. Mauritz C, Schwanke K, Reppel M, et al. Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers. Generation of functional murine cardiac myocytes from induced pluripotent stem cells. Circ Res. 2004;94:952-959. Circulation. 2008;118:507-517. 66. Valiunas V, Doronin S, Valiuniene L, et al. 54. Barbash IM, Chouraqui P, Baron J, et al. Human mesenchymal stem cells make cardiac connexins and form functional gap junctions. Systemic delivery of bone marrow-derived mesenchymal stem cells to the infarcted myocardium: feasibility, cell migration, and body distribution. J Physiol. 2004;555:617-626. Circulation. 2003;108:863-868. 67. Kehat I, Khimovich L, Caspi O, et al. 55. Hou D, Youssef EA, Brinton TJ, et al. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Radiolabeled cell distribution after intramyocardial, intracoronary, and interstitial retrograde coronary venous delivery: implications for current clinical trials. Nat Biotechnol. 2004;22:1282-1289. 68. Xue T, Cho HC, Akar FG, et al. Circulation. 2005;112:I150-I156. Biomimetic approach to cardiac tissue engineering. Functional integration of electrically active cardiac derivatives from genetically engineered human embryonic stem cells with quiescent recipient ventricular cardiomyocytes: insights into the development of cell-based pacemakers. Philos Trans R Soc Lond B Biol Sci. 2007;362:1357-1368. Circulation. 2005;111:11-20. 56. Radisic M, Park H, Gerecht S, Cannizzaro C, Langer R, Vunjak-Novakovic G. 57. Leor J, Gerecht S, Cohen S, et al. 69. Cho HC, Kashiwakura Y, Marban E. Human embryonic stem cell transplantation to repair the infarcted myocardium. Creation of a biological pacemaker by cell fusion. Heart. 2007;93:1278-1284. 70. Rosen MR, Danilo P, Robinson RB. 58. Simpson D, Liu H, Fan TH, Nerem R, Dudley SC Jr. Gene and cell therapy for life-threatening cardiac arrhythmias: will they replace drugs, surgery, and devices? Circ Res. 2007;100:1112-1115. A tissue engineering approach to progenitor cell delivery results in significant cell engraftment and improved myocardial remodeling. Dialogues Cardiovasc Med. 2008;14:44-51. Stem Cells. 2007;25:2350-2357. 71. Edelberg JM, Aird WC, Rosenberg RD. 59. Xiang Z, Liao R, Kelly MS, Spector M. Enhancement of murine cardiac chronotropy by the molecular transfer of the human beta2 adrenergic receptor cDNA. Collagen-GAG scaffolds grafted onto myocardial infarcts in a rat model: a delivery vehicle for mesenchymal stem cells. J Clin Invest. 1998;101:337-343. Tissue Eng. 2006;12:2467-2478. 72. Sakmann B, Trube G. 60. Zimmermann WH, Melnychenko I, Eschenhagen T. Engineered heart tissue for regeneration of diseased hearts. Conductance properties of single inwardly rectifying potassium channels in ventricular cells from guinea-pig heart. Biomaterials. 2004;25:1639-1647. J Physiol. 1984;347:641-657. 24 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Regenerating the heart: gene/cell therapy - Cohen and Gaudette 73. DiFrancesco D. 85. Tse HF, Xue T, Lau CP, et al. The contribution of the “pacemaker” current (If) to generation of spontaneous activity in rabbit sino-atrial node myocytes. Bioartificial sinus node constructed via in vivo gene transfer of an engineered pacemaker HCN channel reduces the dependence on electronic pacemaker in a sick-sinus syndrome model. J Physiol. 1991;434:23-40. Circulation. 2006;114:1000-1011. 74. Miake J, Marban E, Nuss HB. 86. Lieu DK, Chan YC, Lau CP, Tse HF, Siu CW, Li RA. Functional role of inward rectifier current in heart probed by Kir2.1 overexpression and dominant-negative suppression. Overexpression of HCN-encoded pacemaker current silences bioartificial pacemakers. J Clin Invest. 2003;111:1529-1536. Heart Rhythm. 2008;5:1310-1317. 75. Santoro B, Liu DT, Yao H, et al. Identification of a gene encoding a hyperpolarization-activated pacemaker channel of brain. 87. Plotnikov AN, Bucchi A, Shlapakova I, et al. HCN212-channel biological pacemakers manifesting ventricular tachyarrhythmias are responsive to treatment with If blockade. Cell. 1998;93:717-729. Heart Rhythm. 2008;5:282-288. 76. Ludwig A, Zong X, Jeglitsch M, Hofmann F, Biel M. A family of hyperpolarization-activated mammalian cation channels. 88. Altomare C, Terragni B, Brioschi C, et al. Nature. 1998;393:587-591. Heteromeric HCN1-HCN4 channels: a comparison with native pacemaker channels from the rabbit sinoatrial node. 77. DiFrancesco D. J Physiol. 2003;549:347-359. A new interpretation of the pace-maker current in calf Purkinje fibres. 89. Kashiwakura Y, Cho HC, Barth AS, Azene E, Marban E. J Physiol. 1981;314:359-376. Gene transfer of a synthetic pacemaker channel into the heart: a novel strategy for biological pacing. 78. DiFrancesco D, Tortora P. Direct activation of cardiac pacemaker channels by intracellular cyclic AMP. Circulation. 2006;114:1682-1686. Nature. 1991;351:145-147. Biological pacing by implantation of autologous sinoatrial node cells into the canine right ventricle. 90. Zhang H, Shlapakova I, Zhao X, et al. 79. Wainger BJ, DeGennaro M, Santoro B, Siegelbaum SA, Tibbs GR. Circulation. 2008;118:S427-S428. Molecular mechanism of cAMP modulation of HCN pacemaker channels. Nature. 2001;411:805-810. 80. Biel M, Schneider A, Wahl C. Cardiac HCN channels: structure, function, and modulation. Trends Cardiovasc Med. 2002;12:206-212. 81. Qu J, Plotnikov AN, Danilo P Jr, et al. Expression and function of a biological pacemaker in canine heart. Circulation. 2003;107:1106-1109. 82. Plotnikov AN, Shlapakova I, Szabolcs MJ, et al. Xenografted adult human mesenchymal stem cells provide a platform for sustained biological pacemaker function in canine heart. Circulation. 2007;116:706-713. 83. Wang J, Chen S, Siegelbaum SA. Regulation of hyperpolarization-activated HCN channel gating and cAMP modulation due to interactions of COOH terminus and core transmembrane regions. J Gen Physiol. 2001;118:237-250. 84. Bucchi A, Plotnikov AN, Shlapakova I, et al. Wild-type and mutant HCN channels in a tandem biologicalelectronic cardiac pacemaker. Circulation. 2006;114:992-999. 25 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Mending the Broken Heart Expert Answers to Three Key Questions 1 Gene therapy for myocardial infarction–associated congestive heart failure: how far have we got? H. K. Hammond, T. Tang 2 Does cell therapy for myocardial infarction and heart failure work? K. C. Wollert, H. Drexler 3 Gene and cell therapy for life-threatening cardiac arrhythmias: will they replace drugs, surgery, and devices? M. R. Rosen, P. Danilo Jr, R. B. Robinson 27 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated congestive heart failure: how far have we got? H. Kirk Hammond, MD; Tong Tang, PhD VA San Diego Healthcare System - San Diego - Calif - USA Department of Medicine - University of California San Diego - La Jolla - Calif - USA With the advancement of vectors, delivery methods, and newly identified molecular targets, preclinical studies have shown that gene transfer is effective in improving left ventricular contractility and attenuating deleterious left ventricular remodeling in myocardial infarction–associated congestive heart failure (CHF). We are optimistic that these favorable effects will also be seen when tested in patients with CHF associated with myocardial infarction, as well as in patients with CHF from other etiologies. Gene therapy has the potential to be tailored to meet the needs of individual patients. Moreover, when used in conjunction with pharmacological and device management of the patient with CHF, it provides hope for a brighter future for the 23 million patients worldwide with this devastating disease. ongestive heart failure (CHF) is an inexorable disease associated with high morbidity and mortality. CHF has several etiologies, including hypertension, diabetes mellitus, excessive alcohol consumption, valvular heart disease, myocarditis, and familial cardiomyopathy. The majority of cases in developed nations result from coronary artery disease, and subsequent myocardial infarction (MI). Many of these etiologic factors can be altered to avoid the occurrence of CHF altogether—proper treatment of hypertension, surgical intervention for valvular heart disease, risk factor reduction to prevent coronary artery disease, and so forth. C Dialogues Cardiovasc Med. 2009;14:29-36 Once CHF is associated with symptoms at rest or with mild exertion (New York Heart Association [NYHA] class III and IV), 50% of such patients are expected to die within 4 to 5 years — even on optimal medical and device therapy— a prognosis worse than many cancers. Worldwide, it is estimated that 23 million patients have CHF, 1 million new cases are diagnosed yearly, and that every 3 minutes there is another death due to CHF. In the US, CHF is the most common admitting diagnosis in patients >65 years old, in whom there is a 1% incidence. Incidence in this age group is expected to double by 2037. Therefore, despite recent advances, there is an unmet medical need for the millions Copyright © 2009 LLS SAS. All rights reserved 29 Keywords: gene therapy; myocardial infarction; congestive heart failure; angiogenesis; calcium handling; β-adrenergic receptor; adenovirus; adeno-associated virus Address for correspondence: H. Kirk Hammond, MD, VA San Diego Healthcare System (111A), 3350 La Jolla Village Drive, San Diego, CA 92161, USA (e-mail: [email protected]) of patients with CHF. These statistics provide the rationale to develop new, unconventional therapies, such as cell- and gene-based therapies. CARDIAC GENE TRANSFER In general, any gene therapy has three essential components: (i) a vector to carry and express the gene; (ii) a delivery method that safely and efficiently delivers the vector to the organ, tissue, or cell of interest; and (iii) a therapeutic gene tailored to an aspect of the disease being treated. For cardiac gene therapy, there are few suitable vectors, but many candidate genes. The most troublesome component has been the method of delivery. Let us briefly review the most suitable vectors and SELECTED ABBREVIATIONS AND ACRONYMS AAV adeno-associated virus βAR β-adrenergic receptor CHF congestive heart failure FGF fibroblast growth factor GH growth hormone IGF-I insulin-like growth factor–I MI myocardial infarction SERCA2a sarcoendoplasmic reticulum Ca2+-ATPase 2a VEGF vascular endothelial growth factor www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang Lentivirus Adenovirus AAV Figure 1. Virus-mediated gene transfer. The figure depicts the three major virus vectors used for cardiac gene therapy. Once the vector has gained access to the cardiac interstitium, the vectors attach to cell surface receptors or integrins, enter the cell by vesicular transport, traverse the cytoplasm, and enter the nucleus. Adeno-associated virus (AAV) and lentivirus provide persistent transgene expression, as the transgene becomes integrated in the host chromosome. Adenovirus provides extrachromosomal expression. Modified from reference 1: Perkel JM, Slayden C, Swift A. Viral Mediated Gene Delivery. Poster. http://www.sciencemag. org/products/posters/GeneDeli veryPoster.PDF. © 2008 by AAAS/Science. possible delivery methods, before proceeding to a discussion of potential therapeutic genes in MI-associated CHF. Vectors The ideal features of a vector for cardiac gene transfer include: (i) high efficiency; (ii) high specificity (heart and not elsewhere, or cardiac myocyte and not elsewhere, for example); (iii) minimal toxicity or immunogenicity; and (iv) regulated expression. Nonvirus vectors (eg, naked DNA, protein-DNA complexes, and liposomes), despite their easy production, minimal toxicity, and low immunogenicity, have poor transduction efficiency, and their use for CHF gene therapy is limited. Virus vectors have been the main focus for cardiac gene therapy because of their higher myocardial transduction efficiency, and their utility for vascular delivery. The most used virus vectors are adenovirus, adeno-associated virus (AAV), and lentivirus (Figure 1).1 These three vectors, unlike most retrovirus vectors, can be used to obtain gene transfer in cardiac myocytes and other nondividing cells (Table I). Both AAV and lentivirus provide persistent transgene expression, as the transgene becomes integrated in the host chromosome. Adenovirus, in contrast, provides extrachromosomal expression, and is not generally viewed as a longterm expression vector. However, none of these virus vectors are perfect, and extensive efforts have been made to minimize their disadvantages. For example, the deletion of 30 regions of the virus genome encoding proteins specific for the virus per se have provided newer generations of adenovirus with less immunogenicity. In addition, the use of regulated expression vectors enables one to turn expression on or off as needed. Finally, cardiac-specific promoters are available, which limit expression to cardiac myocytes only, even when the vector is more widely disseminated.2 Delivery method Several delivery methods for virus vectors have been used for cardiac gene transfer (Table II). Lentivirus vectors are generally thought to be unsuitable for intracoronary delivery because they do not readily cross the endothelial cell border, and, therefore, direct intramyocardial in- Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang Virus vector classification Chromosomal integration Adenovirus Adeno-associated virus No Yes Advantages Efficient direct intracoronary delivery No risk of insertional mutagenesis Easy to produce Low immunogenicity Vascular delivery may be feasible Permanent expression Lentivirus Yes Very large insert size Permanent expression Disadvantages Immunogenicity Expression not permanent Risk of insertional mutagenesis Difficult to produce Limited insert size Risk of insertional mutagenesis Vascular delivery difficult Difficult to produce Table I. Virus vectors for cardiac gene therapy. jection is a preferred method.3 Intracoronary delivery of adenovirus appears to cause less inflammation in heart compared with other organs, and also circumvents the inflammatory response induced by intramyocardial injection.4,5 However, the intramyocardial injection method is associated with substantial gene transfer efficiency, albeit confined to the areas directly adjacent to the needle tract. Recently, it has been reported that intravenous delivery of AAV vectors—AAV6, AAV8, and AAV9 in particular —may provide substantial gene transfer to heart, lung, and other organs,6 although very high amounts of the virus are required, using doses that may not be feasible in human subjects. Routine intracoronary delivery of AAV is much less efficient than adenovirus.7 Pharmacological agents and mechanical procedures have been used to increase gene transfer efficiency with AAV and adenovirus. Histamine, nitroprusside, serotonin, acetylcholine, sildenafil, vascular endothelial growth factor, and substance P, which increase transcytosis via increases in nitric oxide and cGMP, promote transvascular movement of adenovirus (and perhaps AAV) when delivered by the vascular route — either intracoronary or intravenous.8 AAV and adenovirus vectors can be efficiently delivered to the myocardium using indirect intracoronary delivery, in which the virus vector is injected into the left ventricular (LV) chamber during sustained occlusion of the ascending aorta and proximal pulmonary artery, thus forcing egress into the coronary arteries. Hypothermia, which enables pro- Method longed dwell time without ischemic injury to the heart and brain and simultaneous use of serotonin or acetylcholine, facilitates gene transfer using this method.8,9 Recently, a percutaneous closed-loop recirculation system— essentially a cardiopulmonary bypass without thoracotomy— was shown to provide cardiac gene transfer with AAV— presumably, at least in part, because of the substantially increased dwell time enabled by continuous coronary recirculation of the vector.10 In clinical settings, where safety is the first priority, the gene delivery method that is effective, safe, and easy to apply will prevail as the best choice. Safe methods of delivery will be particularly germane to patients with CHF — invasive methods requiring thoracotomy, cross-clamping of the aorta, or cardiopulmonary bypass will likely be poorly tolerated. GENE THERAPY FOR MI-ASSOCIATED CHF There are three phases in the development of CHF associated with MI, in which a gene therapy may be envisioned: (i) reduction of MI size; (ii) attenuation of LV dilation; and (iii) treatment of fully devel- Advantage Disadvantages Intravenous Simple; relatively noninvasive Low efficiency gene transfer; generalized distribution of vector Myocardial injection Can select region of interest Requires thoracotomy or sophisticated catheter methods; inefficient gene transfer Direct intracoronary Simple; reduced morbidity vs other methods Less efficient than indirect intracoronary Indirect intracoronary Efficient gene transfer Requires pulmonary artery and aortic cross-clamping and thoracotomy or cardiopulmonary bypass Table II. Delivery methods for cardiac gene therapy. 31 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang oped CHF. We will consider the application of gene therapy to these three phases by examining peer-reviewed publications that have used preclinical models of CHF—at this writing (2008), there are no placebocontrolled clinical trials of gene therapy for CHF to review. Reduction of MI size Since acute MI, and consequent loss of LV mass, is the major cause of CHF in developed nations, it is reasonable to ask whether or not gene therapy could reduce infarct size in the setting of acute myocardial infarction, which has been, over the past 25 years, the goal of both thrombolysis and percutaneous coronary interventions. There is overall agreement, however, that reduction of infarct size requires that an occluded vessel be made patent—the earlier, the better—and that a delay of 6 or more hours is generally not associated with substantial reduction in infarct size. With this backdrop, it is difficult to provide a rationale for gene transfer to reduce infarct size in acute MI, given the required delay in vector delivery, transgene expression, and myocardial preserving element (eg, angiogenesis) to result. Indeed, taken together, these requirements of gene therapy, would, in the best circumstances, take many hours to occur. Surprisingly, there have been a few published examples in which gene therapy, administered at the time of MI, or very shortly thereafter, has been associated with a reduction in MI size. Although it seems unlikely that a gene therapy–associated angiogenesis could have occurred rapidly enough to limit infarct size, it is possible that effects related to the inhibition of apoptosis may have contributed. Even so, these possible reductions in MI size are rare, and require experimental coronary occlu- sion with antecedent gene transfer, or vector injection into the myocardium at the time of coronary occlusion—situations that are unlikely to be relevant in clinical acute MI. Attenuation of LV dilation There are several examples in which delivery of genes to the heart, at the time of MI or shortly thereafter, has been associated with a reduction of post-MI LV dilation (adverse remodeling). This is important because, unlike the challenge to reduce infarct size in acute MI— a challenge better achieved with percutaneous coronary interventions or thrombolysis—it is clinically feasible that a gene therapy could be administered after the acute phase of MI, if it were to influence LV remodeling in a favorable manner. Transgenes that have been shown to have this favorable effect on LV remodeling after completed MI include angiogenic genes, antiapoptosis genes, and growth hormone (GH)/insulin-like growth factor-I (IGF-I). Treatment of fully developed CHF Given the abundance of patients with fully developed CHF (23 million worldwide), this phase of treatment may be the most suited to gene therapy, particularly since pharmacological agents have been used so successfully to reduce LV remodeling after MI. On the other hand, one wonders how successful a gene therapy can be when directed to an LV that may have 30% to 40% of its mass already scarred from infarction. Even so, 50% survival has increased from 18 months to 4 to 5 years over the last 25 years with the use of pharmacological agents. It is the hope of those in the field that gene therapy, working on such fundamentally different mechanisms than current medical therapy, may pro- 32 vide better outcomes—giving a patient with severe symptoms the possibility of surviving even longer and feeling better. There are a large number of transgenes that have been used in animal models of CHF with some success, in terms of improved LV function and geometry, exercise capacity, and survival (which will be reviewed in the next section). GENES OF INTEREST WITH SUPPORTIVE PRECLINICAL DATA We have selected promising genes that appear to have favorable influences on the failing heart. Stringent criteria were imposed: (i) convincing demonstration of the presence of CHF; and (ii) gene transfer was performed when CHF was actually present, and not merely given at the time of MI, for example (which may suggest a therapeutic effect but falls short of providing support for a clinically applicable gene therapy per se). Angiogenic and vasculogenic factors The most common cause of CHF in the US and the European Union is MI, and such patients have persistent myocardial ischemia, both in the border zone of the healed infarction, as well as elsewhere in the heart. Even patients with dilated failing hearts unassociated with coronary artery disease have measurable myocardial ischemia. Improving myocardial blood flow by gene transfer of angiogenic/vasculogenic factors is an attractive approach, not only for MI-associated CHF, but also for CHF in general. In pacing-induced CHF in pigs (associated with myocardial ischemia), intracoronary delivery of an adenovirus encoding fibroblast growth factor (FGF)–4, in the setting of ad- Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang vanced CHF, was associated with improvements in global LV function and geometry.11 In the same animal model of CHF, direct intramyocardial injection of plasmid encoding vascular endothelial growth factor (VEGF), showed favorable effects on regional function.12 Finally, a recent paper showed improved regional and global heart function in a pig model of ischemic cardiomyopathy, using intracoronary delivery of an adenovirus encoding FGF5.13 Interestingly, improved LV function was associated not only with the anticipated increase in blood flow in Ad.FGF5-treated animals, but also with increased cardiac myocyte mitosis and reduced apoptosis in the viable ischemic zone. Intramyocardial gene transfer of either hepatocyte growth factor or angiopoietin has been reported to increase regional and global LV function — when delivered at the time of the experimentally induced MI. However, in none of these cases was CHF present at the time of gene transfer, so it remains to be proven that these genes can improve function of the failing heart. β-Adrenergic receptor signaling pathway A hallmark of CHF is impaired β-adrenergic receptor (βAR) signaling, associated with βAR downregulation, Gs/βAR uncoupling (Gs for stimulatory G protein), and abnormalities of adenylyl cyclase (AC) function. These alterations impair cAMP generation, and thus depress myocardial contractility. In the 1980s, treatments for clinical CHF focused on increasing myocardial cAMP levels using pharmacological agents that stimulated the βAR (dobutamine), or decreased the breakdown of cAMP (milrinone). These strategies failed, perhaps owing to sustained high levels of cAMP produc- tion. Current therapy for clinical CHF embraces inhibition, not stimulation of the βAR. Mirroring the poor outcomes seen with βAR-agonist treatments in clinical CHF, are two classic examples from animal studies: (i) chronic infusion of isoproterenol leads to CHF in mice and rats; and (ii) cardiac-directed expression of βAR leads to CHF in transgenic mice. With this backdrop, it is quite surprising that AC, the effector molecule in the βAR signaling pathway, appears to have beneficial effects on the failing heart. AC6 gene expression not only prevents LV hypertrophy and reduces mortality in a genetic model of CHF,14 but also increases LV function, and survival after acute myocardial infarction.15 In addition, AC6 expression prevents further deleterious LV remodeling, and improves cardiac function in MI-associated CHF.16 Why should this element of the βAR signaling pathway— a pathway that governs cAMP production— be so different in its effects compared with strategies designed to stimulate the βAR? Several recent publications have documented that some of the favorable effects of AC6 may not be directly related to increased cAMP generation per se. For example, increased expression of cardiac AC6 corrected calcium-handling defects in cardiomyopathy,14 and, after MI,15 was associated with normalization of cardiac troponin I phosphorylation, and reduced apoptosis in mice with severe CHF.16 AC6 gene transfer influences the phosphorylation and activation of important signaling proteins — independently of cAMP—including inhibition of phospholamban, and activation of Akt.14 These alterations would be predicted to have favorable effects on cardiac function and apoptosis, and shed light on why increased AC6 expression, in contrast to βAR stimula- 33 tion, benefits the failing heart. Gene transfer of AC6 has shown promising results in treating CHF in pigs,17 and a randomized, double-blind, placebo-controlled Phase 1/Phase 2 clinical trial using AC6 gene transfer in patients with stable, but severe CHF has been approved by the Food and Drug Administration, and will be initiated in early 2009. Finally, gene transfer of βARKct,18 an inhibitor of G-protein–coupled receptor kinases, increases contractility in MI-associated CHF in rabbits. These results suggest that controlled increases in βAR signaling, unlike β-agonists and phosphodiesterase inhibitors that lead to unrelenting stimulation of the βAR pathway, may be beneficial for the failing heart. Calcium handling Calcium plays a crucial role in controlling the cardiac contractile process. In every heart beat, calcium is taken up by sarcoplasmic reticulum, through the sarcoendoplasmic reticulum Ca2+-ATPase 2a (SERCA2a) calcium pump, and then released through calcium-release channel ryanodine receptor. Failing hearts exhibit defective calcium uptake and calcium release. Because of the pivotal role of calcium in LV contraction and relaxation, it is a logical focus for gene therapy. Phospholamban is an endogenous muscle-specific inhibitor of SERCA2a, and its phosphorylation at Ser16 releases the inhibition on SERCA2a function. CHF is associated with decreased phospholamban phosphorylation at Ser16. Reduction of phospholamban inhibition of SERCA2a, which promotes the favorable effects of SERCA2a on calcium handling, appears to have favorable effects in genetic cardiomyopathy in mice. To test this ap- Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang proach in fully developed CHF, rats with large infarcts received indirect intracoronary delivery of AAV encoding a pseudophosphorylated mutant phospholamban—a dominant negative mutant that does not inhibit SERCA2a. This resulted in increased cardiac systolic and diastolic function, and reductions in heart weight, cardiac myocyte size, and fibrosis, even 6 months after gene transfer.19 dent.22 Gene transfer of other calcium-handling regulators, for example, protein phosphatase inhibitor 1 and EF-hand calcium-binding proteins sorcin and parvalbumin, has also shown favorable effects on LV function in cardiomyopathic animal models. These methods may be transferrable for CHF treatment, and warrant further studies in MIassociated CHF models. Other candidates There are controversies over whether increased cardiac expression of SERCA2a can improve LV performance in human hearts as it appears to do in rodent hearts, and over how long this effect lasts. However, longterm increases in cardiac SERCA2a content, achieved by AAV-mediated gene transfer, preserved LV systolic function and prevented deleterious LV remodeling in a volume overload–induced CHF model in pigs.20 Two phase 1 clinical studies have been approved that propose to use intracoronary delivery of an AAV encoding SERCA2a to treat patients with CHF. S100A1 belongs to a family of proteins containing 2 EF-hand calciumbinding motifs. S100A1 interacts with SERCA2a, and increases SERCA2a activity. There is also evidence that S100A1 blocks sarcoplasmic reticulum calcium leak during diastole and increases systolic sarcoplasmic reticulum calcium release. Intracoronary delivery of an adenovirus encoding S100A1 restored impaired calcium handling, and improved LV function in myocardial infarction– associated CHF in rats.21 Calcium handling and LV function were restored after intracoronary delivery of an AAV vector with cardiac-specific S100A1 expression into failing rat hearts 10 weeks after MI, where significant calcium-handling impairment, LV dysfunction, and deleterious LV remodeling were evi- Anti-apoptosis During and after acute MI, there is increased apoptosis in myocardium, especially in the border zone of the infarct. Thus, inhibiting apoptosis may attenuate cell loss, thereby preserving myocardial mass. Hepatocyte growth factor and AC6 (discussed previously) appear to reduce apoptosis. Indirect coronary delivery of an adenovirus encoding Bcl-2, an inhibitor of apoptosis, was performed in a rabbit model of regional ischemia-reperfusion injury.23 Bcl-2 expression increased LV ejection fraction and reduced deleterious LV remodeling, which were associated with reduced apoptosis, but the demonstration that this works to treat CHF is lacking. Reduction of reactive oxygen species Myocardial infarction increases reactive oxygen species generation, which has deleterious effects on LV function and geometry. While no studies have conclusively shown that gene therapy with reactive oxygen species scavengers to be effective in fully developed CHF, data in models of myocardial ischemia using delivery of inducible nitric oxide synthase, cyclooxygenase-2, heme oxygenase-1, and extracellular superoxide dismutase are promising.24 Cell cycle reentry Reduced cardiac function after MI is largely caused by cardiac myocyte 34 loss. Although spontaneous cardiac myocyte regeneration may occur after MI, the magnitude of this intrinsic process is insufficient to restore substantial cardiac mass. An intriguing idea is to reinduce adult cardiac myocytes into the cell cycle, and to promote cardiac myocyte regeneration with the hope that this would promote functional recovery after MI. Cardiac-directed expression of cyclin D2 promotes cardiac myocyte proliferation, and new myocardial tissue formation in the infarcted area after MI.25 Associated with these changes were synchronized calcium transients, within the newly regenerated cardiac myocytes and within cardiac myocytes in uninfarcted areas, and increased LV function. Cyclin A2, which is similar to cyclin D2, induces cardiac myocyte proliferation and cardiac regeneration after MI. Presently, these approaches have not yet been used to treat fully developed CHF. GH and IGF-I Although GH (peptide) therapy has been used in clinical CHF, no placebo-controlled, blinded trials have shown efficacy. It is possible that if sustained delivery of GH (or IGF-I) could be achieved—with gene transfer— the approach might be more effective. Although many papers have shown that GH or IGF-I gene therapy—administered at the time of MI— result in improved regional LV function,26 no study has determined whether GH or IGF-I gene therapy is effective in the treatment of fully developed CHF. Given the generally cardiac-friendly effects of GH and IGF-I—both angiogenic and antiapoptotic—they are promising gene therapy candidates. FUTURE DIRECTIONS The pathophysiology of MI-associated CHF is complex. Future studies on molecular signaling pathways will Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang continue to identify new and more effective targets for gene therapy. For example, recent advancement in RNA interference research has identified endogenous small RNAs that specifically silence the expression of target genes. Some of these small RNAs indeed are downregulated after MI. Specific delivery of exogenous small interfering RNA and microRNA into infarcted myocardium presents a new venue for gene therapy. Although there have been several clinical trials of cell-based CHF treatment, cardiac myocyte regeneration has been difficult to document, and the modest clinical benefits reported in these trials may be the result of angiogenesis, which, it can be argued, could be more easily achieved using virus vectors. However, the next phase of gene therapy for CHF will be advanced only when more effective gene transfer vectors and (especially) methods of delivery become available. This work was supported by a Grant-In-Aid from the American Heart Association Western States Affiliate (TT), NIH grants (5P01HL066941; HL081741; HL088426), and a Merit Review Award from the Department of Veteran’s Affairs (HKH). 3. Fleury S, Simeoni E, Zuppinger C, et al. 11. McKirnan M, Lai N, Waldman L, et al. Multiply attenuated, self-inactivating lentiviral vectors efficiently deliver and express genes for extended periods of time in adult rat cardiomyocytes in vivo. Intracoronary gene transfer of fibroblast growth factor-4 increases regional contractile function and responsiveness to adrenergic stimulation in heart failure. Circulation. 2003;107:2375-2382. Card Vasc Regen. 2000;1:11-21. 4. Hammond HK. 12. Leotta E, Patejunas G, Murphy G, et al. Intracoronary gene transfer of fibroblast growth factor in experimental and clinical myocardial ischemia. Gene Ther Regul. 2002;1:325-342. 5. French BA, Mazur W, Geske RS, Bolli R. J Thorac Cardiovasc Surg. 2002;123: 1101-1113. Direct in vivo gene transfer into porcine myocardium using replication-deficient adenoviral vectors. 13. Lynch P, Lee TC, Fallavollita JA, et al. Circulation. 1994;90:2414-2424. 6. Inagaki K, Fuess S, Storm TA, et al. Robust systemic transduction with AAV9 vectors in mice: efficient global cardiac gene transfer superior to that of AAV8. Mol Ther. 2006;14:45-53. 7. Kaspar BK, Roth DM, Lai NC, et al. Myocardial gene transfer and long-term expression following intracoronary delivery of adeno-associated virus. 1. Perkel JM, Slayden C, Swift A. Viral Mediated Gene Delivery. Poster. http://www.sciencemag.org/products/po sters/GeneDeliveryPoster.PDF. Washington, DC: American Association for the Advancement of Science. Accessed December 8, 2008. 2. Su H, Joho S, Huang Y, et al. Adeno-associated viral vector delivers cardiac-specific and hypoxia-inducible VEGF expression in ischemic mouse hearts. Proc Natl Acad Sci U S A. 2004;101: 16280-16285. Intracoronary administration of AdvFGF-5 (fibroblast growth factor-5) ameliorates left ventricular dysfunction and prevents myocyte loss in swine with developing collaterals and ischemic cardiomyopathy. Circulation. 2007;116:I71-I76. 14. Phan HM, Gao MH, Lai NC, et al. New signaling pathways associated with increased cardiac adenylyl cyclase 6 expression: implications for possible congestive heart failure therapy. Trends Cardiovasc Med. 2007;17:215-221. J Gene Med. 2005;7:316-324. 15. Takahashi T, Tang T, Lai NC, et al. 8. Roth DM, Lai NC, Gao MH, et al. Indirect intracoronary delivery of adenovirus encoding adenylyl cyclase increases left ventricular contractile function in mice. REFERENCES Gene therapy with adenovirus-mediated myocardial transfer of vascular endothelial growth factor 121 improves cardiac performance in a pacing model of congestive heart failure. Am J Physiol Heart Circ Physiol. 2004; 287:H172-H177. 9. Hajjar RJ, Schmidt U, Matsui T, et al. Modulation of ventricular function through gene transfer in vivo. Increased cardiac adenylyl cyclase expression is associated with increased survival after myocardial infarction. Circulation. 2006;114:388-396. 16. Lai NC, Tang T, Gao MH, et al. Activation of cardiac adenylyl cyclase expression increases function of the failing ischemic heart in mice. J Am Coll Cardiol. 2008;51:1490-1497. 17. Lai NC, Roth DM, Gao MH, et al. Proc Natl Acad Sci U S A. 1998;95: 5251-5256. Intracoronary adenovirus encoding adenylyl cyclase VI increases left ventricular function in heart failure. 10. Kaye DM, Preovolos A, Marshall T, et al. Circulation. 2004;110:330-336. Percutaneous cardiac recirculation-mediated gene transfer of an inhibitory phospholamban peptide reverses advanced heart failure in large animals. 18. Shah AS, White DC, Emani S, et al. J Am Coll Cardiol. 2007;50:253-260. Circulation. 2001;103:1311-1316. 35 In vivo ventricular gene delivery of a betaadrenergic receptor kinase inhibitor to the failing heart reverses cardiac dysfunction. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene therapy for myocardial infarction–associated CHF - Hammond and Tang 19. Iwanaga Y, Hoshijima M, Gu Y, et al. Chronic phospholamban inhibition prevents progressive cardiac dysfunction and pathological remodeling after infarction in rats. J Clin Invest. 2004;113:727-736. 20. Kawase Y, Ly HQ, Prunier F, et al. Reversal of cardiac dysfunction after longterm expression of SERCA2a by gene transfer in a pre-clinical model of heart failure. J Am Coll Cardiol. 2008;51:1112-1119. 21. Most P, Pleger ST, Volkers M, et al. Cardiac adenoviral S100A1 gene delivery rescues failing myocardium. J Clin Invest. 2004;114:1550-1563. 22. Pleger ST, Most P, Boucher M, et al. Stable myocardial-specific AAV6-S100A1 gene therapy results in chronic functional heart failure rescue. Circulation. 2007;115:2506-2515. 23. Chatterjee S, Stewart AS, Bish LT, et al. Viral gene transfer of the antiapoptotic factor Bcl-2 protects against chronic postischemic heart failure. Circulation. 2002;106:I212-I217. 24. Bolli R, Li QH, Tang XL, et al. The late phase of preconditioning and its natural clinical application—gene therapy. Heart Fail Rev. 2007;12:189-199. 25. Hassink RJ, Pasumarthi KB, Nakajima H, et al. Cardiomyocyte cell cycle activation improves cardiac function after myocardial infarction. Cardiovasc Res. 2008;78:18-25. 26. Chao W, Matsui T, Novikov MS, et al. Strategic advantages of insulin-like growth factor-I expression for cardioprotection. J Gene Med. 2003;5:277-286. 36 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? Kai C. Wollert, MD; Helmut Drexler, MD Department of Cardiology and Angiology - Hannover Medical School - Hannover - GERMANY The infarcted heart heals by scar formation, and large myocardial infarctions typically result in heart failure. Although adult stem cells with the capacity to transform into various cardiac cell types and to secrete cardioprotective cytokines have been identified, endogenous repair mechanisms in the adult heart are not sufficient for meaningful tissue regeneration. These observations, however, suggest that it may be feasible to develop interventions aimed at enhancing these processes, and to promote functional and, eventually, structural recovery of the infarcted heart. Recent randomized clinical trials indicate that intracoronary delivery of bone marrow (stem) cells leads to an improvement in systolic function after myocardial infarction, thereby providing the first evidence that cell therapy may work in patients. Keywords: myocardial infarction; heart failure; stem cell therapy; transdifferentiation; paracrine factors Address for correspondence: Kai C. Wollert, MD, Professor of Cardiology, Dept of Cardiology and Angiology, Hannover Medical School, Carl-NeubergStr. 1, 30625 Hannover, Germany (e-mail: [email protected]) Dialogues Cardiovasc Med. 2009;14:37-43 Copyright © 2009 LLS SAS. All rights reserved odern reperfusion strategies and advances in pharmacological management have resulted in an increasing proportion of patients surviving after an acute myocardial infarction (AMI). The resulting loss of viable myocardium sets the stage for progressive left ventricular remodeling in many patients. The extent of cardiac remodeling after an AMI is closely related to the size of the infarct: larger infarcts result in a greater extent of left ventricular remodeling and a worse prognosis. As a consequence, AMI has become the most common cause of heart failure in many countries. However, none of our current therapies addresses the underlying cause of the remodeling process, ie, the critical loss of myocardium in the infarcted area. M THE CONCEPT OF CELL THERAPY It has recently been observed that cardiomyocytes in the infarct border zone may reenter the cell cycle and divide after an AMI.1 In addition, genetic fate–mapping studies support the notion that endogenous stem cells may be a source of new cardiomyocytes after ischemic injury.2 Moreover, there are data to suggest that the injured myocardium attracts circulating stem cells and progenitor cells that may positively affect the healing response and functional recovery after an AMI via the release of paracrine factors.3 The overall capacity of the adult 37 heart for regeneration is limited, however, and the vast majority of necrotic cardiomyocytes are replaced by scar tissue after an AMI. Still, the existence of endogenous regenerative mechanisms may open the way to mimicking and amplifying these processes therapeutically. Two main approaches to achieve myocardial tissue replacement and functional enhancement have been envisioned: the use of isolated cells delivered directly to the diseased myocardium (cell therapy—the focus of the present paper), or the use of a combination of cells and biomaterials to generate functional three-dimensional tissues in vitro before implanting them into the body (tissue engineering). As the heart is composed of 30% cardiomyocytes and 70% nonmyocytes, such as endothelial cells, smooth muscle cells, and fibroblasts, cardiac regeneration is not only a matter of cardiac myocyte addition, but also of nonmyocyte supplementation. POTENTIAL DONOR CELLS There are two different cell sources that might be used for cell transplantation: autologous and allogeneic cells. Autologous cells are obtained from the patients themselves, and pose no risk of immune rejection. However, the functional activities of autologous cells may be negatively affected by underlying cardiovascular risk factors and dis- www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler ease.4 Transplantation of most allogeneic cells will require immunosuppressive therapy to avoid immunological reactions. Experimentally, differentiated cells as well as stem and progenitor cells have been employed for cell therapy. Each cell type has its own profile of advan- Cell types tages, limitations, and practicability issues, and may have an impact on cardiac structure and/or function through distinct mechanisms (Table I). In general, differentiated cells show a lower proliferation rate and a diminished survival rate after transplantation when compared with Pros stem and progenitor cells.5 Stem cells are capable of self-renewal, transformation into dedicated progenitor cells, and differentiation into specialized progeny. Depending on their differentiation potential, stem cells are classified as being pluripotent (capable of differenti- Cons Cardiomyocytes Fetal or neonatal cardiac myocytes - Integrate electrically and mechanically with host myocardium - Limited supply - Ethical and legal issues - Need for use in an allogeneic setting - Limited capacity for ex vivo expansion - Poor survival after transplantation Cells with no apparent capacity for cardiomyocyte differentiation Skeletal myoblasts - Easy to obtain (skeletal muscle biopsy) - Ex vivo expansion requires a culturing process lasting several days - Retain skeletal muscle phenotype after intracardiac transplantation Mesenchymal stem cells - Can be expanded ex vivo - Promote paracrine effects - Low immunogenicity (can possibly be used in an allogeneic setting) - Ex vivo expansion requires a culturing process lasting several days - Transdifferentiation into mature cardiomyocytes uncertain Endothelial progenitor cells - Easy to obtain from blood or bone marrow - Promote paracrine effects - Some may differentiate into endothelial cells - Enhance neovascularization - Transdifferentiation into cardiomyocytes uncertain Unfractionated bone marrow cells - Easy to obtain - Contain several stem- and progenitorcell populations - Promote paracrine effects - Low percentage of stem and progenitor cells - Probably no meaningful transdifferentiation into cardiomyocytes Cardiomyocyte progenitor cells Embryonic stem (ES) - Pluripotent cells with capacity for cells differentiation into vascular cells and cardiomyocytes - Can be expanded and differentiated into cardiomyocytes ex vivo prior to transplantation - Risk of teratoma formation if contaminating pluripotent cells are transplanted - Ethical and legal issues - Need for use in an allogeneic setting Induced pluripotent stem (iPS) cells - Same as for ES cells - Can be used in an autologous setting - Risk of teratoma formation if contaminating pluripotent cells are transplanted - Risk of insertional mutagenesis Spermatogonial stem cells - Same as for ES cells - Can be used in an autologous setting - Risk of teratoma formation if contaminating pluripotent cells are transplanted - Can be obtained only from Cardiac resident progenitor cells - Allegedly multipotent cells with capacity for differentiation into vascular cells and cardiomyocytes - Can be expanded ex vivo - Specific cell surface markers, and protocols for cell isolation and expansion need to be better defined Table I. Cell types used for cell therapy and tissue engineering. 38 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler Cell delivery strategies Pros Cons Intravenous infusion - Least invasive - Cell trapping in the lungs and other tissues - Limited cell delivery to the heart Intracoronary infusion - Homogeneous cell delivery to the site of injury - Cells that are retained will have adequate blood supply - Open infarct-related artery required - Limited cell retention in infarcted area - Not suitable for delivery of large cells which may cause microembolization (eg, skeletal myoblasts, mesenchymal stem cells) Transendocardial injection - Electromechanical mapping of the endocardial surface can be used to delineate viable, ischemic, and scarred myocardium before cell injections - Creates islands of cells with limited blood supply and poor cell survival - May not be safe in patients with acute myocardial infarction (AMI), when cells are injected in friable necrotic tissue Transepicardial injection - Allows for direct visualization of the myocardium, and a targeted application of cells to scarred areas and/or the border zone of an infarct - Creates islands of cells with limited blood supply and poor survival - Open heart surgery required - Invasiveness hampers its use as a stand-alone therapy, or its use in the setting of AMI Table II. Cell delivery strategies. ating into any of the three germ layers) or multipotent (giving rise to a limited number of other cell types). CELL TRANSPLANTATION STRATEGIES The goals of any cell delivery strategy are to transplant sufficient numbers of cells into the myocardial region of interest and to achieve maximum retention of cells within that area (Table II). Cell retention may be defined as the fraction of transplanted cells retained in the myocardium for a short period of time. The local milieu is an important determinant of cell retention, as it will influence short-term cell survival and, if a transvascular approach is used, cell adhesion, transmigration through the vascular wall, and tissue invasion. Transvascular strategies are especially suited to the treatment of recently infarcted and reperfused myocardium, when chemoattractants are highly expressed.6 Selective intracoronary application delivers a maximum concentration of cells homogeneously to the site of injury. Unselected bone marrow cells have been delivered via the intracoronary route in patients after AMI. In these studies, cells were delivered through the central lumen of an over-the-wire balloon catheter during transient balloon inflations to maximize the contact time of the cells with the microcirculation of the infarct-related artery. In experimental models, intravenous delivery of endothelial progenitors cells and mesenchymal stem cells has been shown to improve cardiac function after AMI. However, cell homing to noncardiac organs limits the applicability of this approach. Indeed, in a recent clinical study, homing of unselected bone marrow cells to the infarct region was observed only after intracoronary stopflow delivery, but not after intravenous infusion.7 Direct injection techniques may be more appropriate for patients presenting late in the disease process, when an occluded coronary artery precludes transvascular cell delivery or when homing signals are expressed at low levels in the heart (scar tissue). 39 CURRENT STATUS OF CELL THERAPY IN PATIENTS WITH ACUTE MYOCARDIAL INFARCTION AND HEART FAILURE Preclinical studies have shown that transplantation of bone marrow–derived hematopoietic stem cells, endothelial progenitor cells, or mesenchymal stem cells can promote functional improvements in animal models of AMI. Transdifferentiation of the transplanted cells into cardiomyocytes and endothelial cells has been offered as an explanation, but the quantitative importance of stable cell engraftment and transdifferentiation for the functional effects has been challenged. It is now believed that the reported improvements in these models are mediated predominantly by paracrine effects (Figure 1, page 40)3,8-13 Clinicians welcomed these animal studies with great enthusiasm, and, fairly rapidly, clinical trials were designed to translate these finding into the clinical scenarios of post-AMI or heart failure patients. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler Cell homing and tissue integration Paracrine effects EC differentiation SMC differentiation Vasculogenesis Cardiac differentiation and fusion Angiogenesis Activation of CPCs Arteriogenesis Cardiomyocyte proliferation Cardiomyocyte apoptosis Modulation of inflammation Cardiomyogenesis Scar remodeling Functional improvements Randomized trials using unselected bone marrow cells Most clinical investigators have chosen a pragmatic approach by using unfractionated bone marrow cells, which contain different stem and progenitor cell populations, as well as more differentiated hematopoietic cell types. In all of these studies, the cells were delivered into the reperfused and stented infarctrelated artery by using a stop-flow balloon catheter approach. In four trials, cells were delivered a few days after coronary reperfusion to enhance systolic function and prevent adverse remodeling; in one trial, bone marrow cells were transplanted in patients with ischemic heart failure, months or years after an AMI (Table III)14-18,23 The combined experiences from these studies indicate that intracoronary delivery of unselected bone marrow cells is feasible and probably safe. The outcome of these randomized trials has been mixed, however, with some studies showing significant improve- ments in global and regional left ventricular systolic function,14,15,18 one trial showing improvements in regional function only,16 and one trial reporting no significant improvements at all.18 While the reasons for these heterogeneous results are difficult to resolve, it has been argued that differences in cell preparation methods and in timing of cell transfer may have been critical.15-17,19 Recent meta-analyses of published randomized and nonrandomized studies, involving a total of approximately 1000 patients, support the notion that bone marrow cell transfer contributes to modest improvements in cardiac function after AMI, above and beyond current interventional and medical therapy.20-22 Skeletal myoblast transplantation Recently, the first randomized, placebo-controlled study of skeletal myoblast transplantation after myocardial infarction has been published.23 Patients were treated with culture- 40 Figure 1. Proposed mechanisms of action of cell therapy after myocardial infarction. The relative contributions, for example, of paracrine effects vs differentiation events, will depend on the transplanted cell type(s) and on the microenvironment of the host tissue. Abbreviations: CPC, resident cardiac progenitor cell; EC, endothelial cell; SMC, smooth muscle cell. Adapted from reference 13: Dimmeler S, Burchfield J, Zeiher AM. Cell-based therapy of myocardial infarction. Arterioscler Thromb Vasc Biol. 2008;28:208-216. Copyright © 2008, Lippincott Williams & Wilkins. expanded, autologous skeletal myoblasts or placebo, at least 4 weeks after AMI. Cells were injected into the infarct border zone during bypass surgery. Myoblast transplantation did not improve regional or global left ventricular function, the primary end points of the trial. Notably, however, a significant decrease in left ventricular volumes was noted after cell therapy. A greater incidence of arrhythmias was noted in the myoblast-treated patients, but this did not translate into differences in major adverse cardiac events after 6 months. ISSUES TO ADDRESS AT BENCH AND BEDSIDE The mixed results from the randomized bone marrow cell trials remind us that procedural issues, such as the cell preparation method, cell dosage, and timing of cell transfer need to be further refined in upcoming studies. Different cell populations and cell delivery methods may be required depending on Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler Outcome Time of cell delivery Dose (after AMI) Improved No change Design n* Cell type BOOST14 Open, controlled 30 treated 30 controls Nucleated BMCs 128 mL 6±1 days Global LVEF LVEDV REPAIR-AMI15 Placebocontrolled 95 treated 92 controls Mononucleated BMCs 50 mL 3-6 days Global LVEF LVEDV Leuven-AMI16 Placebocontrolled 32 treated 34 controls Mononucleated BMCs 130 mL 1 day ASTAMI17 Open, controlled 47 treated 50 controls Lymphocytic BMCs 50 mL 6±1 days – Global LVEF, LVEDV TOPCARE-CHD18 Open, controlled 35 treated 23 controls Mononucleated BMCs 50 mL 81±72 months Global LVEF LVEDV MAGIC†23 Placebocontrolled 67 treated 30 controls Skeletal myoblasts 400 800 106 >4 weeks LVEDV, LVESV Regional contractility, global LVEF Study Regional Global LVEF, contractility LVEDV *Only patients with complete imaging studies are considered. † Dose refers to the average amount of bone marrow that was harvested, or the number of transplanted skeletal myoblasts. Table III. Randomized cell therapy trials in patients with acute myocardial infarction or ischemic heart failure. In BOOST, cells were prepared by gelatine polysuccinate density gradient sedimentation, which retrieves all nucleated cell types from the bone marrow. REPAIR-AMI, TOPCARE-CHD, and Leuven-AMI employed a Ficoll gradient, which recovers the mononuclear cell fraction. Although a similar cell isolation protocol was used in ASTAMI, the cell yield was lower as compared with REPAIR-AMI. Remodeling was assessed by MRI in Leuven-AMI, BOOST, and in ASTAMI, by left ventricular angiography in REPAIR-AMI and TOPCARE-CHD, and by echocardiography in MAGIC. Abbreviations: AMI, acute myocardial infarction; BMC, bone marrow cell; LVEDV, left ventricular end-diastolic volume; LVEF, left ventricular ejection fraction; n, number of patients; LVESV, left ventricular end-systolic volume. Study acronyms: ASTAMI, Autologous Stem-cell Transplantation in Acute Myocardial Infarction; BOOST, BOne marrOw transfer to enhance ST-elevation infarct regeneration; Leuven-AMI, Leuven-Acute Myocardial Infarction; MAGIC, Myoblast Autologous Grafting in Ischemic Cardiomyopathy; REPAIR-AMI, Reinfusion of Enriched Progenitor cells And Infarct Remodeling in Acute Myocardial Infarction; TOPCARE-CHD, Transplantation Of Progenitor Cells And REcovery of LV [left ventricular] function in patients with Chronic ischemic Heart Disease. whether the AMI occurred recently or not to achieve optimum therapeutic effects early or late after AMI. Patient subgroups that derive the greatest benefit from cell transfer need to be identified prospectively, eg, patients presenting late after symptom onset in whom little myocardial salvage can be expected from reperfusion therapy. The impact of bone marrow cell transfer on clinical end points is currently unknown. Although a significant reduction in the combined end point of death, myocardial infarction, or revascularization was observed in one trial,24 no such effects were observed in other (smaller) trials.25 Ultimately, large outcome trials using optimized cell preparation and delivery strategies need to be performed. Cell-labeling studies indicate that less than 5% of unselected, nucleated bone marrow cells are retained in the infarcted area after intracoronary delivery in patients.7 It is conceivable that pharmacologic strategies might be used to enhance the homing capacity or other functional parameters of the cells; experimental studies are pointing in this direction.26,27 The ultimate goal of stem cell therapy is to replace the infarcted area with new contractile and fully integrated cardiomyocytes. While unselected bone marrow cells may have a favorable impact on systolic function, they probably do not make new myocardium. The lack of true cardiac regeneration should stimulate further basic research into the 41 therapeutic prospects of cardiomyocyte progenitor cells. Recent data support the idea that paracrine effects play an important role in patients undergoing bone marrow cell transfer. In fact, bone marrow cells have been shown to secrete a number of proangiogenic factors,28 consistent with the improvements in microvascular function observed after bone marrow cell transfer in patients.29 Experimental studies suggest that enhanced angiogenesis after AMI may improve infarct healing and energy metabolism in the infarct border zone. Further investigation of candidate factors in animal models may ultimately enable more specific and powerful therapeutic strategies after AMI. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler REFERENCES 1. Beltrami AP, Urbanek K, Kajstura J, et al. 10. Murry CE, Soonpaa MH, Reinecke H, et al. 18. Assmus B, Honold J, Schachinger V, et al. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Transcoronary transplantation of progenitor cells after myocardial infarction. Evidence that human cardiac myocytes divide after myocardial infarction. Nature. 2004;428:664-668. N Engl J Med. 2001;344:1750-1757. 11. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. 2. Hsieh PC, Segers VF, Davis ME, et al. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Cardioprotective c-kit+ cells are from the bone marrow and regulate the myocardial balance of angiogenic cytokines. J Clin Invest. 2006;116:1865-1877. 4. Dimmeler S, Leri A. Aging and disease as modifiers of efficacy of cell therapy. Circ Res. 2008;102:1319-1330. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature. 2004;428:668-673. Eur Heart J. 2007;28:766-772. 12. Kinnaird T, Stabile E, Burnett MS, et al. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ Res. 2004;94:678-685. Impact of intracoronary cell therapy on left ventricular function in the setting of acute myocardial infarction: a collaborative systematic review and meta-analysis of controlled clinical trials. 14. Wollert KC, Meyer GP, Lotz J, et al. Circulation. 2008;118:1425-1432. 7. Hofmann M, Wollert KC, Meyer GP, et al. Monitoring of bone marrow cell homing into the infarcted human myocardium. Circulation. 2005;111:2198-2202. 8. Dai W, Hale SL, Martin BJ, et al. Allogeneic mesenchymal stem cell transplantation in postinfarcted rat myocardium: short- and long-term effects. Circulation. 2005;112:214-223. Arch Intern Med. 2007;167:989-997. Cell-based therapy of myocardial infarction. Stem-cell-based therapy and lessons from the heart. Pilot trial on determinants of progenitor cell recruitment to the infarcted human myocardium. Adult bone marrow-derived cells for cardiac repair: a systematic review and meta-analysis. 21. Lipinski MJ, Biondi-Zoccai GG, Abbate A, et al. Arterioscler Thromb Vasc Biol. 2008; 28:208-216. 6. Schachinger V, Aicher A, Dobert N, et al. 20. Abdel-Latif A, Bolli R, Tleyjeh IM, et al. 13. Dimmeler S, Burchfield J, Zeiher AM. 5. Passier R, van Laake LW, Mummery CL. Nature. 2008;453:322-329. 19. Seeger FH, Tonn T, Krzossok N, Zeiher AM, Dimmeler S. Cell isolation procedures matter: a comparison of different isolation protocols of bone marrow mononuclear cells used for cell therapy in patients with acute myocardial infarction. Nat Med. 2007;13:970-974. 3. Fazel S, Cimini M, Chen L, et al. N Engl J Med. 2006;355:1222-1232. Intracoronary autologous bone-marrow cell transfer after myocardial infarction: the BOOST randomised controlled clinical trial. Lancet. 2004;364:141-148. 15. Schachinger V, Erbs S, Elsasser A, et al. Intracoronary bone marrow-derived progenitor cells in acute myocardial infarction. N Engl J Med. 2006;355:1210-1221. 16. Janssens S, Dubois C, Bogaert J, et al. Autologous bone marrow-derived stem-cell transfer in patients with ST-segment elevation myocardial infarction: double-blind, randomised controlled trial. Lancet. 2006;367:113-121. J Am Coll Cardiol. 2007;50:1761-1767. 22. Burt RK, Loh Y, Pearce W, et al. Clinical applications of blood-derived and marrow-derived stem cells for nonmalignant diseases. JAMA. 2008;299:925-936. 23. Menasche P, Alfieri O, Janssens S, et al. The Myoblast Autologous Grafting in Ischemic Cardiomyopathy (MAGIC) trial: first randomized placebo-controlled study of myoblast transplantation. Circulation. 2008;117:1189-1200. 24. Schachinger V, Erbs S, Elsasser A, et al. 9. Ziegelhoeffer T, Fernandez B, Kostin S, et al. 17. Lunde K, Solheim S, Aakhus S, et al. Bone marrow-derived cells do not incorporate into the adult growing vasculature. Intracoronary injection of mononuclear bone marrow cells in acute myocardial infarction. Improved clinical outcome after intracoronary administration of bone-marrow-derived progenitor cells in acute myocardial infarction: final 1-year results of the REPAIR-AMI trial. Circ Res. 2004;94:230-238. N Engl J Med. 2006;355:1199-1209. Eur Heart J. 2006;27:2775-2783. 42 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Does cell therapy for myocardial infarction and heart failure work? - Wollert and Drexler 25. Arnesen H, Lunde K, Aakhus S, Forfang K. Cell therapy in myocardial infarction. Lancet. 2007;369:2142-2143. 26. Sasaki K, Heeschen C, Aicher A, et al. Ex vivo pretreatment of bone marrow mononuclear cells with endothelial NO synthase enhancer AVE9488 enhances their functional activity for cell therapy. Proc Natl Acad Sci U S A. 2006;103: 14537-14541. 27. Sorrentino SA, Bahlmann FH, Besler C, et al. Oxidant stress impairs in vivo reendothelialization capacity of endothelial progenitor cells from patients with type 2 diabetes mellitus: restoration by the peroxisome proliferator-activated receptor-gamma agonist rosiglitazone. Circulation. 2007;116:163-173. 28. Korf-Klingebiel M, Kempf T, Sauer T, et al. Bone marrow cells are a rich source of growth factors and cytokines: implications for cell therapy trials after myocardial infarction. Eur Heart J. 2008;29:2851-2858. 29. Erbs S, Linke A, Schachinger V, et al. Restoration of microvascular function in the infarct-related artery by intracoronary transplantation of bone marrow progenitor cells in patients with acute myocardial infarction: the Doppler Substudy of the Reinfusion of Enriched Progenitor Cells and Infarct Remodeling in Acute Myocardial Infarction (REPAIR-AMI) trial. Circulation. 2007;116:366-374. 43 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy for life-threatening cardiac arrhythmias: will they replace drugs, surgery, and devices? Michael R. Rosen, MD; Peter Danilo Jr, PhD; Richard B. Robinson, PhD Center for Molecular Therapeutics - Columbia University - New York, NY - USA Ventricular tachycardia and/or fibrillation result in 200 000 400 000 sudden cardiac deaths each year in the US alone and atrial fibrillation currently afflicting up to 2.3 million Americans each year. Gene and cell therapies for cardiac arrhythmias are nascent fields whose raisons d’être derive from: (i) the problematic state of arrhythmia treatment today (especially atrial and ventricular tachyarrhythmias for which drugs, devices, and ablation remain more stopgap measures than optimal interventions); and, (ii) the opportunity to learn, and potentially treat and cure these arrhythmias, by exploring new technologies. Our review examines the state of antiarrhythmic therapy today and the new directions being taken. V entricular tachycardia (VT) and/or ventricular fibrillation (VF) lead to 200 000 to 400 000 sudden cardiac deaths each year in the US.1 Atrial fibrillation (AF) today afflicts about 2.3 million Americans, and may reach 12 to 16 million by 2050.2 Reentry accounts for nearly all AF and approximately 85% of arrhythmias in ischemic heart disease (IHD).3 In one conceptualization, the reentrant waveform follows a well-defined anatomic pathway, reaching its point of origin after the standing wave that is the action potential has ended, and the tissue at that site is no longer refractory. For this to happen, a number of changes in conduction must occur, including unidirectional block and slow retrograde propagation.4,5 Another concept is functional reentry,6 in which the length and/or position of the reentrant path changes over time. Dialogues Cardiovasc Med. 2009;14:44-51 Therapeutic approaches to reentry use drugs to prolong the effective refractory period (ERP), and/or to slow/block conduction, and/or surgery or catheter ablation to interrupt the pathway. For example, in AF, flecainide, dofetilide, sotalol, and amiodarone are representative drugs having therapeutic roles.7 Regrettably, clinical trials have demonstrated virtually all antiarrhythmics tested to be proarrhythmic.8 A dif- Copyright © 2009 LLS SAS. All rights reserved 44 Keywords: ventricular tachycardia, ventricular fibrillation, atrial fibrillation, reentry, myocardial infarction, stem cells, viral vectors Address for correspondence: Michael R. Rosen, MD, Gustavus A. Pfeiffer Professor of Pharmacology, Professor of Pediatrics, Director, Center for Molecular Therapeutics, Columbia University, PH 7W-321, 630 West 168 Street, New York, NY, 10032, USA (e-mail: [email protected]) ferent pharmacological approach uses angiotensin-converting enzyme inhibitors or angiotensin II type 1 (AT1) receptor blockers to reduce paroxysmal AF recurrences.9 Radiofrequency ablation terminates AF caused by triggered foci in pulmonary veins prevents recurrences, and appears more effective than antiarrhythmic drugs. However, ablation manifests variable success rates: recurrence is more frequent than anticipated. No antiarrhythmic drug is acceptable primary or sole therapy for VT/VF in the setting of IHD.8 In contrast, several clinical device trials8 have shown survival benefit in treating potentially lethal arrhythmias SELECTED ABBREVIATIONS AND ACRONYMS AF atrial fibrillation AV atrioventricular ERP effective refractory period hERG human ether-a-go-go gene hMSCs human mesenchymal stem cells IHD ischemic heart disease SkM1 skeletal muscle Na channel VF ventricular fibrillation VT ventricular tachycardia www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others in patients with IHD. But device and combined drug/device approaches to VT/VF carry a high economic price. Emotional and physical tolls comprise a different and important cost. All told, new approaches to prevention/termination would be welcome. A B C D E F NOVEL APPROACHES TO PREVENTING/ TERMINATING REENTRY Nearly 100 years ago, Mines4,5 conceptualized and experimentally demonstrated reentry and communicated its potential role in clinical arrhythmias. He postulated that the relationship between path length, propagation velocity, and refractoriness determines whether reentry will evolve and persist. Much subsequent discovery derives from Mines’ observations: in the process we have learned of the multiple wavelet and leading circle concepts of reentry, of anisotropy, and of rotors. The actions of clinically-used IKr and INa-blocking antiarrhythmic drugs dovetail with Mines’ concepts: they prolong ERP and slow conduction to interrupt reentry.10 Yet Mines’ concepts avail us of much more information that has not been applied to antiarrhythmic advantage. Figure 1 (from Schmitt and Erlanger’s 1928 adaptation of Mines5) explores this: a waveform activates the peripheral conducting system, but if there is a region of depressed conduction (grey), antegrade propagation may fail (Panels A-C) and reentry may occur (Panels D-E). We can terminate the reentrant loop by further depressing conduction until it is blocked bidirectionally and/or by prolonging ERP. Both are outcomes of antiarrhythmic drug therapy: the conundrum in prolonging ERP is that we usually prolong repolarization as well and encounter proarrhythmia.10 We can cut the Figure 1. Cardiac arrhythmias and reentry concepts. (Panels A-E) Schmitt and Erlanger’s5 concept of reentry sees an impulse propagating through the conducting system, with antegrade conduction blocked in one limb, and then conducting retrogradely through to reactivate the proximal portion of the system. (Panel F) shows the antegrade activation that would be expected to occur if one could facilitate conduction through depressed regions. loop surgically or using catheter ablation, in the process perhaps creating a scar that may then serve as a nidus for further reentry. Mines recognized therapeutic options other than prolonging ERP or blocking conduction.4,5 We should be able to speed conduction such that a waveform “catches its tail,” thereby encountering refractory tissue and failing to propagate further. 45 We also should be able to prolong ERP relative to repolarization, but without prolonging repolarization itself.10 To date, neither approach has achieved consistent success. Moreover, a clear derivative of Mines’ model is the concept of regional therapies that might modify a portion of a pathway. For example if the Figure 1 pathway were made to propagate normally (Panel F), then reentry would not commence. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others CURRENT STATUS OF ANTIARRHYTHMIC GENE AND CELL THERAPIES Antiarrhythmic gene and cell therapies have: (i) resulted in biological pacemakers for treating heart block11; (ii) utilized regional delivery of viral constructs for treating VT or AF12; and (iii) developed alternatives to surgery for ablating the atrioventricular (AV) junction in AF.13-15 General approaches to gene and cell therapy are summarized in Figure 2. To modify cardiac rhythm directly, we can use gene therapy carried broblasts and the human mesenchymal stem cells (hMSCs) in Figure 2. Finally (though not depicted), administration of plasmids in the absence of a carrier has shown poor incorporation, although electroporation is being attempted as a means to improve this approach. The other antiarrhythmic approach (Figure 2, right) is to repair/regenerate myocardium using embryonic or hMSCs or more refined precursor cells (reviewed elsewhere in this issue). Reduction of arrhythmias is an expected secondary outcome of this strategy. Modify cardiac function Modify cardiac structure/function Deliver a gene Regenerate/repair tissue • hESCs • hMSCs • Cardiac precursor cells hMSC expression • Electroporation • Viral expression Figure 2. Approaches to cell and gene therapy of arrhythmias. To modify cardiac function directly, we can use gene or cell therapy (left). The other option is to use cardiac repair/regeneration strategies (right). See text for discussion. Abbreviations: AA, adeno-associated virus; hESC, human embryonic stem cell; hMSC, human mesenchymal stem cell. via viral vectors. These include: (i) adenovirus, which expresses briefly and episomally, and is used largely for proof-of-concept experiments; (ii) adeno-associated virus, which shows long persistence in expression, but whose genomic incorporation is uncertain; and (iii) lentivirus, which results in genomic incorporation. Despite persistent questions regarding long-term impact on human subjects, these viruses are being used in clinical trials. Cells loaded via electroporation or viral vectors have also been used to carry gene constructs. These include fi- The treatment of tachyarrhythmias has been more challenging than that of bradyarrhythmias. Issues include design of therapeutic constructs and whether to administer treatment globally or locally, as discussed below. Global versus local administration Cell and gene therapy of arrhythmias Viral expression • Adenovirus • AAV • Lentivirus into a pacemaker line,16 adult hMSCs used as platforms to carry pacemaker genes17,18 and fibroblasts used to carry pacemaker genes to myocytes with which they have been fused.19 Finally, in AV block cell-engineered bypass tracts have carried sinus impulses to the ventricles.20 The leading edge of gene/cell arrhythmia research has focused on bradyarrhythmias and creation of biological pacemakers or AV bridges. Biological pacing strategies have included overexpressing β2-adrenergic receptors, transfecting a dominant negative construct to reduce IK1, overexpressing the hyperpolarization-activated cyclic nucleotidegated (HCN) gene family to increase pacemaker current and using mutagenesis to create designer pacemakers based on HCN or K channel genes.11 Cell therapy has seen human embryonic stem cells coaxed 46 Permeabilizing agents, vasodilators, and vascular endothelial growth factor (VEGF) have been used to facilitate gene delivery to large or localized regions of the heart.21,22 Cooling and aortic cross-clamping have been employed to improve gene delivery, either through the coronary artery or by the flooding of a chamber or chambers.21,22 Not only do these approaches appear excessive for clinical application, but the best success to date has seen about 50% of cells in any region transfected, with viral transfer being diffusion-limited and especially problematic in the ventricles.22 Tempering interest in some viral vectors are concerns about inflammation, chronic illness, or neoplasia. These issues led us to explore hMSCs as platforms for gene delivery. It is exciting that hMSCs can be loaded with specific gene constructs17,18 and delivered to the heart without eliciting inflammation or rejection, and without differentiating into other cell types.23 However, the long-term stability of hMSC therapies raises concern (eg, migration to other sites, differentiation into other cell types, and duration of Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others expression of genes of interest).11 The use of various markers to trace cell location should facilitate our understanding of the extent of hMSC localization to sites of administration.11 Hence, viral vector–based therapies have not yet been applied clinically to arrhythmia management, but have been effective in proof-of-concept experiments (see below), suggesting that gene therapy can be of use. Cell therapies, generally, have been intended to regenerate and repair myocardium rather than to be specifically antiarrhythmic. While we have found hMSCs to be adequate delivery platforms for ion channel generated currents, we have only followed them for 6 weeks.18 The question of long-term applicability will have to await long-term studies of hMSC survival as well as comparison with genomically-incorporated viral constructs. Novel ion channel constructs as antiarrhythmic interventions Given the feasibility of gene/cell therapy approaches, a potential advantage is that, unlike drugs, they do not limit us to the channels and transporters expressed by native cardiac myocytes. Instead, channels resident in other tissues, or manmade mutant or chimeric channels with more favorable biophysical properties can be employed. Such a unique arsenal of antiarrhythmic tools allows a “rational” approach to antiarrhythmic therapy, in which the biophysical properties of an ideal therapeutic agent are defined, synthesized, and delivered. A general approach to administering gene therapy constructs is suggested in Figure 3. Theoretically, we might employ novel Na channels or connexins to speed conduction, and/or alter the properties of inward or repolar- Extracellular Cell 1: inside Cell 2: inside Extracellular 0 mV Normal cell Depolarized cell Figure 3. Gene therapy and ion channel constructs. (Top) Two myocytes and their ion channels. Sodium (Na) and potassium (K) channels are transmembrane structures that respectively carry Na current inward (red arrow), resulting in depolarization, and K current outward (yellow arrow), resulting in repolarization. Connexins (Cx) populate the intercellular gap junctions, and permit the flow of electrons from one cell to another, facilitating the propagation of the cardiac impulse. (Bottom) Schematic of action potentials from normal and depolarized (infarcted) ventricular myocytes. The upper traces are the action potentials, and the lower traces are the maximal upstroke velocity of phase 0 (Vmax ) of the action potential (reflecting rapid Na entry). Note how the depolarized cell has a low Vmax , and how this results in slowed propagation. • • Created using Servier technology (Servier Medical Art: http://www.servier.com/Smart/ ImageBank.aspx?id=729), with permission. Action potentials obtained, with permission, from: Lue WM, Boyden PA. Circulation. 1992;85: 1175-1188. Copyright © 2008, American Heart Association Inc. izing currents to produce postrepolarization refractoriness. Alternatively, we might deliver small interfering RNA to target specific channels for inhibition and to block conduction at localized sites. Atrial fibrillation A major goal of gene therapy experiments on atrial fibrillation has been to induce atrioventricular block. To this end, G-α i2 overexpression via AV nodal artery injection in pig was used to suppress basal adenylyl cyclase activity and, via amplified vagal tone, to indirectly reduce the Ca current.13 During sinus rhythm, AV conduction slowed and ERP was prolonged, and, during AF, there was a 20% reduction in 47 ventricular rate.13 Other strategies reported are the creation of an AV nodal site of Ca channel blockade,14 or the implantation of fibroblasts to induce AV nodal scarring and block.15 All these approaches exemplify local gene delivery whose therapeutic intent is to produce rate control. Whether they will become a practical alternative to radiofrequency ablation is uncertain. Another experimental AF therapy aimed at rhythm control uses an ion channel mutation Q9E-hMiRP1 (a contributor to long QT syndrome induced by IKr-blocking drugs). Levy et al administered the construct into the atrial epicardium of pigs: about 15% of cells manifested up- Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others Membrane potential/channel m gate h gate –90 mV/SCN5A –60 mV/SCN5A –60 mV/SkM1 Figure 4. Comparison of SCN5A and SkM1 sodium (Na) channels as inward charge carriers in normal and depolarized cells (the latter mimicking an epicardial border zone in myocardial infarct). Depicted are the sequence of channel states from resting, through open (depolarized), to repolarizing. The Na channel is shown as are the “m” and “h” gates (in open or closed configurations). (Top) At –90 mV, SCN5A (and SkM1) carry the Na current inward (yellow arrow) in their open channel state. (Middle) At –60 mV, the h gate of the SCN5A channel is closed, and there is no flow of current inward. (Bottom) By contrast, at the same membrane potential (–60 mV), there is an inward flow of Na current through the SkM1 channel (yellow arrow). Hence, SkM1 is capable of carrying robust current at membrane potentials at which SCN5A is inactive and nonfunctional. Created using Servier technology (Servier Medical Art: http://www.servier. com/Smart/ ImageBank.aspx?id=729), with permission. Channel state Resting Depolarized take. Clarithromycin was then infused and profoundly blocked IKr . This led the investigators to hypothesize that, in AF, they might achieve regional atrial IKr blockade without prolonging the QT interval.24 Ventricular tachycardia/fibrillation Whereas myocardial infarct–induced arrhythmias might respond to local therapy, variations in anatomy from patient to patient require extensive mapping to determine sites at which to localize therapy. For example, Reddy et al demonstrated that mapping to identify sites for local radiofrequency ablation reduced the need for defibrillation in patients who had devices implanted for secondary prevention.25 Using mapping to identify the border zone of an infarct in a canine model, we have replaced ablation with intramyocardially administered gene therapy in preliminary studies and—without destroying tissue—achieved a reduction in VT/VF incidence.26 Repolarized Specific gene therapies for ischemic arrhythmias • Speeding conduction via Na channels or connexins At least 10 different Na channel genes encode α-subunits in the mammalian genome, and these have been cloned from brain, spinal cord, skeletal and cardiac muscle, uterus, and glia.6 Since slow conduction is an essential feature of reentrant cardiac arrhythmias, we sought other mammalian Na channels that might have more favorable properties than the cardiac Na channel in circumstances that favor slow conduction (Figure 4).26 One such circumstance is membrane depolarization, as in myocardial infarction (Figure 3). Here, the voltage dependence of steady state Na channel inactivation is of interest. The midpoint of the cardiac Na channel (SCN5A) is negative to –73 mV. This is important because, in infarcted tissue when myocytes are depolarized to –65 mV, virtually all SCN5A- 48 derived cardiac Na channels are inactivated. In contrast, skeletal muscle (SkM1) Na channels have an inactivation midpoint of –68 mV, and almost half of these channels would be available to open during an action potential in a depolarized cell. This suggests that Na channels with more favorable biophysical properties than SCN5A, such as SkM1, might be a useful antiarrhythmic therapy (Figure 4). Data from our laboratory have demonstrated the effectiveness of this approach in a canine model, in which the incidence of inducible polymorphic VT was 75% of controls and 17% of SkM1-administered dogs 5 days postinfarction.26 Moreover, as shown in Figure 5, SkM1 administration reduced electrogram fragmentation and increased Vmax of phase 0 (consistent with more rapid conduction), as had been predicted for SkM1. Several studies have lent credence to the importance of connexins and, hence, gap junctions in arrhythmias. For example, overexpression of Cx45 results in ventricular tachycardia in mice,27 while mutations of Cx40 are associated with atrial fibrillation in humans.28 Studies of the epicardial border zone of heal• Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others ing canine myocardial infarcts have demonstrated altered connexin distribution and density in regions important to the generation of reentrant ventricular tachycardia.29 The modulation of gap junctions as an antiarrhythmic strategy initially attempted to block conduction. However, the gap junctional blockers used to date have not been channelspecific or isoform-specific, and, in disrupting coupling between cells, have been found to cause potentially fatal arrhythmias. On the positive side, antiarrhythmic peptides have been used to increase junctional conductance. One such pep- tide, rotigaptide, appears to target Cx43 specifically,30 and is purportedly antiarrhythmic. • Targeting diastolic membrane potential In VT in the setting of a partially healed infarct, the viable, but depolarized, tissue in the border zone provides the substrate for a reentrant arrhythmia (Figure 3).6 A logical approach to enhance conduction in these circumstances is to hyperpolarize diastolic membrane potential, thereby making more Na current available. In normal myocytes, the diastolic membrane po- A ECG 1 mV 3 EG 2 3 2 L 4 200 ms 1 4 B 0 1 1 40 mV 200 V/s 2 40 mV 200 V/s 100 ms 100 ms tential is largely set by the inward rectifier IK1 (generated by Kir2.1 with some contribution from Kir2.2).31 Studies on the overexpression of these channels are in progress. • Enhancing rate responsiveness and/or refractoriness Reentrant arrhythmias require reexcitation of tissue by a propagating waveform. Here, an intervention that facilitates recovery of excitability in the pathway may restore antegrade activation and forestall retrograde invasion of that path by the reentering waveform. Alternatively, it may speed propagation of the reentering waveform so that it encounters tissue that remains refractory. Hua et al32 showed that 6-fold overexpression of native human ether-a-go-go gene (hERG) eliminates T-wave alternans in isolated canine ventricular myocytes and in computer simulations. Using a different approach, Sasano et al delivered a dominant negative hERG mutant (HERG-G628S) via vascular infusion to a peri-infarct zone of pigs.12 Monomorphic VT had been consistently inducible in infarcted animals before gene transfer, but, 1 week later, all HERG-G628S-transferred pigs showed no such arrhythmia. This result emphasizes the therapeutic potential of a different local approach to VT therapy in chronic infarcts. CONCLUSIONS Figure 5. Effect of SkM1 adenoviral infection of epicardial site. A canine heart was infarcted and injected with SkM1+green fluorescent protein (GFP) adenovirus 1 week before this experiment. (A) Photo of the left ventricular epicardial surface. Each panel (1 to 4) in the photograph displays a surface ECG (top) and a bipolar local electrogram (bottom). The broken line demarcates the infarcted myocardium (bottom) from the noninfarcted myocardium (top). Note how the local electrogram of the noninjected infarcted site 2 is markedly fragmented. Infarcted site 1 (injected with SkM1) has a normal electrogram as do noninfarcted sites 3 and 4. (B) Hematoxylin and eosin stains of tissues from sites 1 (+SkM1) and 2 (no SkM1) show infarcted myocardium ( 200). Inset in site 1 is GFP positive; that in site 2 is GFP negative ( 400). Representative action potential recorded from site 1 has higher Vmax and amplitude than that from site 2. • Modified after reference 26, with permission: Lau DH, Clausen C, Sosunov EA, et al. Epicardial border zone overexpression of skeletal muscle sodium channel, SkM1, normalizes activation, preserves conduction and suppresses ventricular arrhythmia: an in silico, in vivo, in vitro study. Circulation. 2009;119:19-27. Copyright © 2009, American Heart Association. 49 How best to prevent/treat arrhythmias that confer significant morbidity and/or are life-threatening is a question that 40 years of targeted pharmacologic therapy and many more years of empirical drug therapy have not answered. While surgery, ablation, and cardioverter-defibrillators are newer, robust alternatives, all appear draconian when compared with a targeted therapy that can be administered via catheter Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others and doesn’t destroy tissue. Gene and cell therapies meld principles for rational drug design with a new approach to treatment. The approach starts by simulating an optimal change in an ion current present in the heart, and then selects constructs independent of their tissue origins based on their close mimicry of the proposed optimal change. Additionally, use of novel genes not normally found in heart expands our therapeutic universe. Also innovative are the focus on regional delivery of these novel ion channel constructs, the use of hMSCs as platforms for therapeutic intervention, and the harnessing of these new therapies to mechanistically test old, but to date largely untestable, concepts (eg, speed conduction, increase ERP/repolarization ratio without prolonging repolarization). These approaches should not be interpreted as providing a “quick fix.” However, the ability to prepare constructs and to apply them based on an understanding of arrhythmogenic mechanisms makes it highly likely that: (i) definitive answers— whether positive or negative—will be obtained to the questions we have regarding improvement of antiarrhythmic therapy; and (ii) we will understand why, mechanistically, an approach has succeeded, failed, or, as a worst case, been proarrhythmic. Negative answers will be as important as positive: of signal importance is that as accurately and rapidly as possible we find the proper route, vector and/or platform, and construct for reducing the threat to the population of the arrhythmias of concern. The authors gratefully acknowledge the assistance of Eileen Franey in the preparation of this manuscript. Research was supported in part by USPHS-NHLBI grants HL-28958 and HL-67101. REFERENCES 1. Myerburg RJ, Feigal DW Jr, Lindsay BD. Life-threatening malfunction of implantable cardiac devices. N Engl J Med. 2006;354:2309-2311. 2. Miyasaka Y, Barnes ME, Gersh BJ, et al. 8. Spooner PM, Rosen MR. Perspectives on arrhythmogenesis, antiarrhythmic strategies and sudden cardiac death. In: Spooner PM, Rosen MR, eds. Foundations of Cardiac Arrhythmias. New York, NY; Marcel Dekker Inc; 2000: 1-20. 9. Ducharme A, Swedberg K, Pfeffer MA, et al. Circulation. 2006;114:119-125. CHARM Investigators. Prevention of atrial fibrillation in patients with symptomatic chronic heart failure by candesartan in the Candesartan in Heart failure: Assessment of Reduction in Mortality and morbidity (CHARM) program. 3. Wit AL, Janse MJ. Am Heart J. 2006;152:86-92. Experimental models of ventricular tachycardia and fibrillation caused by ischemia and infarction. 10. Members of the Sicilian Gambit. Secular trends in incidence of atrial fibrillation in Olmsted County, Minnesota, 1980 to 2000, and implications on the projections for future prevalence. Circulation. 1992:85:I32-I142. Antiarrhythmic Therapy: A Pathophysiologic Approach. Armonk, NY: Futura Publishing Co; 1994. 4. Mines GR. 11. Rosen M. On circulating excitations in heart muscles and their possible relation to tachycardia and fibrillation. Biological pacemaking: in our lifetime? Heart Rhythm. 2005;2:418-428. Trans R Soc Can. 1914;IV:43-52. 12. Sasano T, McDonald AD, Kikuchi K, Donahue JK. 5. Schmitt FO, Erlanger J. Molecular ablation of ventricular tachycardia after myocardial infarction. Directional differences in the conduction of the impulse through heart muscle and their possible relations to extrasystolic and fibrillary contractions. Am J Physiol. 1928;87:326-347. 6. Allessie MA, Bonke FIM, Schopman FJG. Nature Med. 2006;12:1256-1258. 13. Bauer A, McDonald AD, Nasir K, et al. Inhibitory G protein overexpression provides physiologically relevant heart rate control in persistent atrial fibrillation. Circus movement in rabbit atrial muscle as a mechanism of tachycardia. III. The “leading circle” concept: a new model of circus movement in cardiac tissue without the involvement of an anatomical obstacle. Circulation. 2004;110:3115-3120. Circ Res. 1977;41:9-18. Circ Res. 2004;95:398-405. 7. Fuster V, Ryden LE, Cannom DS, et al. 15. Bunch TJ, Mahapatra S, Bruce GK, et al. American College of Cardiology. American Heart Association Task Force on Practice Guidelines. European Society of Cardiology Committee for Practice Guidelines. ACC/ AHA/ESC 2006 guidelines for the management of patients with atrial fibrillation– executive summary: a report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and the European Society of Cardiology Committee for Practice Guidelines. Impact of transforming growth factor-β1 on atrioventricular node conduction modification by injected autologous fibroblasts in the canine heart. J Am Coll Cardiol. 2006;48:854-906. Nature Biotechnol. 2004;22:1282-1289. 50 14. Murata M, Cingolani E, McDonald AD, Donahue JK, Marban E. Creation of a genetic calcium channel blocker by targeted gem gene transfer in the heart. Circulation. 2006;113:2485-2494. 16. Kehat I, Khimovich L, Caspi O, et al. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Gene and cell therapy of arrhythmias - Rosen and others 17. Potapova I, Plotnikov A, Lu Z, et al. Human mesenchymal stem cell as a gene delivery system to create cardiac pacemakers. Circ Res. 2004;94:841-959. 18. Plotnikov AP, Shlapakova I, Szabolcs MJ, et al. Xenografted adult human mesenchymal stem cells provide a platform for sustained biological pacemaker function in canine heart. Circulation. 2007;116:706-713. 19. Cho HC, Kashiwakura Y, Marban E. Creation of a biological pacemaker by cell fusion. 26. Lau DH, Clausen C, Sosunov EA, et al. Epicardial border zone overexpression of skeletal muscle sodium channel, SkM1, normalizes activation, preserves conduction and suppresses ventricular arrhythmia: an in silico, in vivo, in vitro study. Circulation. 2009;119:19-27. 27. Betsuyaku T, Nnebe NS, Sundset R, Patibandla S, Krueger CM, Yamada KA. Overexpression of cardiac connexin45 increases susceptibility to ventricular tachyarrhythmias in vivo. Circ Res. 2007;100:1112-1115. Am J Physiol Heart Circ Physiol. 2006;290:H163-H171. 20. Choi YH, Stamm C, Hammer PE, et al. 28. Gollob MH, Jones DL, Krahn AD, et al. Cardiac conduction through engineered tissue. Somatic mutations in the connexin 40 gene (GJA5) in atrial fibrillation. Am J Pathol. 2006;169:72-85. N Engl J Med. 2006;354:2677-2688. 21. Lehnart SE, Donahue JK. Coronary perfusion cocktails for in vivo gene transfer. Methods Mol Biol. 2003;219:213-218. 22. Roth DM, Lai NC, Gao MH, et al. Indirect intracoronary delivery of adenovirus encoding adenylyl cyclase increases left ventricular contractile function in mice. Am J Physiol Heart Circ Physiol. 2004;287:H172-H177. 23. Zimmett JM, Hare JM. Emerging role for bone marrow derived mesenchymal stem cells in myocardial regenerative therapy. 29. Peters NS, Coromilas J, Severs NJ, Wit AL. Disturbed connexin43 gap junction distribution correlates with the location of reentrant circuits in the epicardial border zone of healing canine infarcts that cause ventricular tachycardia. Circulation. 1997;95:988-996. 30. Dhein S, Larsen BD, Petersen JS, Mohr FW. Effects of the new antiarrhythmic peptide ZP123 on epicardial activation and repolarization pattern. Cell Commun Adhes. 2003;10:371-378. Basic Res Cardiol. 2005;100:471-481. 31. Zaritsky JJ, Redell JB, Tempel BL, Schwarz TL. 24. Perlstein I, Burton DY, Ryan K, et al. The consequences of disrupting cardiac inwardly rectifying K+ current (IK1) as revealed by the targeted deletion of the murine Kir2.1 and Kir2.2 genes. Posttranslational control of a cardiac ion channel transgene in vivo: clarithromycinhMiRP1-Q9E interactions. J Physiol. 2001;533:697-710. Hum Gene Ther. 2005;16:906-910. 32. Hua F, Johns DC, Gilmore RF Jr. 25. Reddy VY, Reynolds MR, Neuzil P, et al. Prophylactic catheter ablation for the prevention of defibrillator therapy. N Engl J Med. 2007;357:2657-2665. Suppression of electrical alternans by overexpression of HERG in canine ventricular myocytes. Am J Physiol Heart Circ Physiol. 2004;286:H2342-H2352. 51 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Matters @ Heart Doppler and his principle Richard J. Bing, MD Professor of Medicine - USC (Em) - Director of Experimental Cardiology - Huntington Medical Research Institute Visiting Professor in Chemistry - California Institute of Technology - Calif - USA e encounter the name of Doppler when we listen to the weather forecast, or read about the beginning of the universe (the Big Bang, the red shift), or when we study diagnostic reports on heart disease. Who was this man who gave us the tools to explore these divergent subjects? W Christian Doppler was born in November 1803 in Salzburg, Austria, and died in Venice, Italy, on March 17, 1853. He came from a family of master stonemasons and showed exceptional gifts for this craft. But because of his poor health, his father considered him suitable for the bookkeeping function of the family business. But it soon became apparent that Doppler had outstanding talents in mathematics. He was sent to the Polytechnic School in Vienna, but he disliked the instructions and called it “a one-sided education.” At the age of 21, he returned to his native Salzburg and finished his edu- Christian Doppler. Contemporary portrait. Courtesy of the Christian Doppler Foundation, Salzburg, Austria. cation while he supported himself by giving classes in mathematics and physics. He then returned for four years to the Polytechnic in Vienna. Like other great scientists, including Einstein, Doppler received many rejections in response to applications for positions, and finally was compelled to become a bookkeeper. Discouraged, he set his eye on America and in Munich, he discussed with the American consul the possibilities of finding a teaching position in America. He even sold his possessions to finance his journey. But while in Munich, he received two offers to teach in high schools in Switzerland or in Prague, which was then part of the Austrian Empire. These were not university positions, but involved high school teaching. Doppler chose the position in Prague, where he again encountered opposition and frustration, making him anxious to leave Prague. Like Einstein who failed to attain an appointment at the techni- Adapted from: Richard J. Bing, MD. Past Truth and Present Poetry: Medical Discoveries and the People Behind Them. First published as a series in Heart News and Views (News Bulletin of the International Society for Heart Research). The essays are now available as a collection published under the same title in 2006, by TFM PUBLISHING LTD Nr. Shrewsbury ISBN 1903378443. By kind permission of Heart News and Views (www.ishrworld.org) and TFM PUBLISHING LTD (www.tfmpublishing.com). Address for correspondence: Richard J. Bing, MD, Director, Experimental Cardiology, Em, HMRI - Huntington Medical Research Institute, 99 N Cl Molino Avenue, Pasadena, CA 91011, USA (e-mail: [email protected]) Dialogues Cardiovasc Med. 2009;14:53-56 Copyright © 2009 LLS SAS. All rights reserved View of Salzburg with the Hohensalzburg fortress, the Cathedral (Dom), and Peterskirche. Oil on canvas, 19th-century. Private collection. Photo © Bonhams, London, UK/The Bridgeman Art Library. 53 www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Doppler and his principle - Bing cal high school in Zurich, Doppler had an unsuccessful interview for a position at the Polytechnic Institute in Vienna. Interviews with other institutions were equally unsuccessful. In Prague, he published a new optical instrument, called the distometer, for measuring distances and discussed the aberrations of light and sound in a rotating medium. It was at a meeting of the Natural Sciences Section in Prague that Christian Doppler postulated the theory that immortalized his name. He and his family left Prague after he was appointed Professor of Mathematics in a small Czechoslovakian town. Finally, he was elected a Full Member of the Imperial Academy of Science in Vienna and awarded an honorary doctorate from the philosophical faculty. He was then appointed to join the Polytechnic Institute in Vienna and finally, was authorized to found an institute of physics at the Imperial University in Vienna. There, a 20-yearold Augustinian monk, Johann Gregor Mendel, took a written and oral examination to study at the University A The former Imperial-Royal Polytechnic Institute (k. k. Polytechnisches Institut) in Vienna. Now Vienna University of Technology. © Clemens Pfeiffer, with kind permission. of Vienna, but Doppler was not very impressed by his mathematical ability and Mendel was refused admission to the university. Mendel was finally admitted. He later laid the foundation of genetics as an abbot in a monastery. Doppler suffered from tuberculosis, Low-pitched siren High-pitched siren B Red shift Blue shift Sketch illustrating the Doppler Principle. A. Doppler effect with sound: when the fire engine recedes from the observer, the apparent wavelength increases, the apparent frequency decreases, and the sound is therefore lowpitched. Conversely, when the fired engine approaches, the sound is perceived as high-pitched by the observer. B. Doppler effect with light: light emitted from a galaxy receding from the observer will show a lengthening of its wavelength, hence a “red-shift,” whereas a galaxy approaching the observer will exhibit a “blue shift”: the fact that galaxies consistently show a red shift has been taken as proof that our universe is expanding. 54 Statue of Gregor Mendel in Augustinian monastery of St Thomas in Brno (Czech Republic). It was in the monastery’s gardens that Mendel discovered the laws of genetic inheritance by observing the transmission of traits in pea plants. © Zarco, with kind permission. which had spread to the larynx and made speaking increasingly difficult. In 1852, his health had so deteriorated that he took a six-month period of convalescence in Venice, where the climate was supposed to influence the course of tuberculosis. He died in 1853 and was buried in Venice. Aside from being plagued by ill health all his life, Doppler was continuously afraid of losing his livelihood, with good reasons as his career demonstrates. His friends called him modest, thrifty, and correct. Like many physicists before and after him, he derived his inspiration for his principle from observations of natural phenomena. As he wrote, “We know from general experience that a ship of moderately deep draught which is steering toward the oncoming waves has to receive, in the same period of time, more waves and with a greater impact than one which is not moving or is even moving along in the direction of the waves. If this is valid for the waves of water, then why should it not also be applied with necessary modification to air and ether waves?” Doppler applied his principle first to astronomy. In his article on the “Col- Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Doppler and his principle - Bing The Doppler Effect. Contemporary oil painting (2000) by Alan Kingsbury, featuring the horns, hornplayer, and the train in the experiment that confirmed Doppler’s Principle. Private collection. © The Bridgeman Art Library. ored light of double stars and other constellations of the heavens” he argued that all stars emitted white light and that the color of some of the stars was due to their motion toward us or away from us. Actually this was an erroneous conclusion because on the approach of a star only a slight shift would be produced, but no change in color would take place. But the principle is correct; an apparent shift in the frequency of waves received by an observer depends on the relative motion between the observer and the source of the waves. This principle was immediately attacked and as recently as 1965 objections against his principle were published. Others opposed Doppler’s theory because of its simplicity. One of them, Petzval, used the argument that, “Without the application of differential equations, it is not possible to enter the realms of great science.” Obviously, his critics thought that great truth could not be found in a few lines and through an equation with only one unknown; at least one differential equation is necessary. In 1845, a Dutch scientist, Buys Ballot of Utrecht, confirmed Doppler’s principle on the railway between Utrecht and Amsterdam. Using a locomotive capable of attaining the at that time incredible speed of 40 mph, to pull an open cart in which horn players were riding, Ballot attempted to observe changes in the apparent pitch of the notes played by the musicians as they approached or receded. However, the experiment was performed in February and the musicians had trouble blowing their instruments because of the cold, and the project was postponed; in June of that year, the validity of Doppler’s theory was finally confirmed. In cardiology, the principle was first utilized to detect cardiac motion and time the opening and closing of the cardiac valves. Satomura used a continuous ultrasound beam transmitted through the chest wall to the heart which, reflected from the heart structures underwent a frequency shift, or Doppler shift, of the transmitted sound; its magnitude and direction were based on the speed and direction of movements of the heart. The frequency of the reflected sound was proportional to the velocity of components of the target. Once the velocity of blood flow in the aorta could be recorded, the technique was adapted for measurement of cardiac output. The most important development in cardiac Doppler analysis was the introduction of pulse wave Doppler, which allows localization of flow velocity measurements to specific valves and chambers. It is based on the principle that blood flow in a small area within the heart can be recorded by the use of intermittent pulses of transmitted sound. The receiver then listens for reflected Left-ventricular and aortic pulsed Doppler echocardiographic flow patterns through esophageal wall in a normal adult subject. Abbreviations: AML, anterior mitral leaflet; AV, aortic valve; FLOW, spectral recording indicating quality and direction of flow at position of sample volume; LA, left atrium; LV, left ventricle; RVOT, right-ventricular outflow tract; SV, sample volume. Reproduced from: Hisanaga K, Hisanaga A, Ichie Y, et al. Transoesophageal pulsed Doppler echocardiography. Lancet. 1979;1:53-54. © 1979, Elsevier Ltd. 55 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Doppler and his principle - Bing FURTHER READING Bing R, ed. Abstract painting? Color-coded Doppler ultrasound duplex scanning image of carotid vessels (arterial blood red, venous blood blue). © Wellcome Photo Library/ Wellcome Images. sound only at the end of the time interval required for the pulse to travel from the transducer to the area of interest and back. This way it is possible to localize murmur, determine orifice size from jet diameter, and measure pulmonary flow and pulmonary artery pressure. Doppler’s principle has made it possible to determine the ejection fraction of the heart, one of the most valuable measurements in cardiology. It has to a large extent, together with echocardiography, replaced cardiac catheterization, particularly in children with congenital heart disease. The correlation between measured Doppler flow velocities and pressure gradients form the basis for assessment of valvular and vascular stenosis, prosthetic valves, and permitted estimation of chamber pressure. Simultaneous determination of velocity in several areas of the heart can also be performed. The digitally processed system displays velocity by color-coding. Current instrumentation allows for superimposition of colorcoded velocity on the tomography image. Doppler’s principle is also applicable to the diagnosis of congenital malformations of the heart in utero. Christian Doppler’s life shows again that scientific accomplishments do not guarantee personal happiness. One of the main reasons in Doppler’s case was ill health. Since his early youth, Doppler was plagued by progressive respiratory disease, due to tuberculosis, which involved the larynx. In addition, Doppler’s concept was new. Like many great discoveries, it was simple and it was direct. It was just this simplicity and directness, which caused other scientists of limited outlook to suspect his principle. Lesser scientists often judge a new discovery by its complexity, which they find attractive. Furthermore, Doppler’s discovery had no practical applicability. It took more than 100 years to make an impact on cosmology, meteorology, and medicine. As Einstein has said, “Ach der Mensch betrügt sich gern, nimmt die Schale für den Kern,” or translated freely “How man does fool himself, mistaking the outer shell for the inner truth.” 56 Cardiology: The Evolution of the Science and the Art, 2nd Ed. New Brunswick, NJ: Rutgers University Press; 1999. Eden A. Christian Doppler, Thinker and Benefactor. Salzburg, Austria: Christian Doppler Institute for Medical Science and Technology; 1988. Franklin DL, Ellis RM, Rushmer RF. Aortic blood flow in dogs during treadmill exercise. J Appl Phys. 1959;14:809-812. Gillispie CC, ed. Christian Doppler, in: Dictionary of Scientific Biography. New York, NY: Charles Scribner’s Sons; 1971:4:167. Hisanaga K, Hisanaga A, Ichie Y, et al. Transoesophageal pulsed Doppler echocardiography. Lancet. 1979;1:53-54. Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Mending the Broken Heart Summaries of Ten Seminal Papers David H. Lau, MD, PhD Columbia University College of Physicians and Surgeons - Division of Cardiology - Department of Medicine New York, NY - USA (e-mail: [email protected]) Dialogues Cardiovasc Med. 2009;14:57-67 1 6 Evidence that human cardiac myocytes divide after myocardial infarction Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers A. P. Beltrami and others. N Engl J Med. 2001 I. Potapova and others. Cir Res. 2004 2 7 Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes Regenerating the heart M. A. Laflamme and C. E. Murry. Nat Biotechnol. 2005 I. Kehat and others. J Clin Invest. 2001 3 8 Biological pacemaker created by gene transfer Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction J. Miake and others. Nature. 2002 D. L. Kraitchman and others. Circulation. 2005 4 9 Heart regeneration in zebrafish Molecular ablation of ventricular tachycardia after myocardial infarction K. D. Poss and others. Science. 2002 T. Sasano and others. Nat Med. 2006 5 10 Adult cardiac stem cells are multipotent and support myocardial regeneration Theoretical impact of the injection of material into the myocardium: a finite element model simulation A. P. Beltrami and others. Cell. 2003 S. T. Wall and others. Circulation. 2006 Selection of seminal papers by David H. Lau, MD, PhD Columbia University College of Physicians and Surgeons - Division of Cardiology - Department of Medicine - 622 West 168th Street, PH-347 New York, NY 10032 - USA Copyright © 2009 LLS SAS. All rights reserved Highlights of the years by Ian Mudway, MD Lung Biology - Division of Life Sciences Franklin Williams Building 150 Stamford Street - London SE1 9NN - UK 57 www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Evidence that human cardiac myocytes divide after myocardial infarction A. P. Beltrami, K. Urbanek, J. Kajstura, S. M. Yan, N. Finato, R. Bussani, B. Nadal-Ginard, F. Silvestri, A. Leri, C. A. Beltrami, P. Anversa N Engl J Med. 2001;344:1750-1757 R emarkable examples of regeneration can be found throughout nature. Newts regrow whole limbs. A flatworm can form a complete flatworm from a small portion of itself. Regeneration of damaged tissue is essential for long human life as our bodies mend fractured bone, repair torn muscle and skin, and renew blood cells. It was long held as dogma that the adult human brain and heart were not capable of regenerating new neurons or cardiomyocytes. This belief was underscored by clinical experience. After a stroke, infarcted brain tissue appears permanently lost as demonstrated by decades of experience with brain imaging. In the heart, myocardial infarction (MI) is a common early insult leading to impaired cardiac function and clinical heart failure. Scar tissue replaces the infarcted region, not new myocardium. In the 1990s, breakthrough discoveries in hippocampal biology challenged the dogma that adult brains cannot regenerate. To revisit the regenerative potential of the adult heart, Beltrami et al conducted a thorough painstaking examination of human hearts with recent MI to search for evidence of mitosis. They found strong evidence that cell division does indeed occur in adult human hearts. similar numbers of cardiomyocytes with visible evidence of mitosis were present (70-fold increase in infarct border and 24-fold increase in distant myocardium). To definitely prove completion of mitosis and formation of two daughter cells would require labeling studies in patients, unlikely with current technology. The results provide strong evidence that the heart’s response to injury is cardiomyocyte proliferation to compensate for the lost cells. The presence of Ki-67/α-actin stained cells with mitotic spindles in the normal controls suggests that there maybe a continuous turnover of cardiomyocytes throughout life. As with all paradigm-shifting discoveries, more questions than answers were raised. What are the molecular signals that govern proliferation? What is the origin of the dividing cell? Are they differentiated cardiomyocytes that reenter the cell cycle? Are there resident cardiac stem cells? Do extracardiac stem cells home to the heart and proliferate into cardiomyocytes? Is regenerating activity in the infarct border zone a substrate for post-MI arrhythmias? It is clear that cardiac proliferation after a myocardial infarction is not a clinically meaningful process. However, this exciting research area will yield insights that may change tomorrow’s treatment of heart failure. Thirteen hearts from patients who died 4 to 12 days after suffering a MI were harvested 7 to 17 hours after death. Samples were taken from the infarct border zone and a site distant to the infarct. Standard histological methods were used and sections were analyzed with confocal microscopy for the presence of Ki-67, a nucleolus component found in every cell cycle phase except G0 (resting state), and α-sarcomeric actin, expressed only by cardiomyocytes. Therefore, any cell staining positive to both Ki-67 and α-actin is a cardiomyocyte undergoing the process of cell division. Over 100 000 nuclei were analyzed in the infarct border and the normal region in each heart. The results are startling and provide much food for thought. In the infarct border, there was an 84-fold greater number of double labeled Ki-67/α-actin cells than in the comparable control region. When the site distant from the infarct was compared, a 28 times greater number of Ki-67/α-actin cells was seen. Using an antitubulin antibody to identify mitotic spindles, 2001/1901 The US stock market crashes for the first time; Wilhelm Conrad Röntgen, the discoverer of x-rays, is awarded the Nobel Prize in Physics; and Queen Victoria, Queen of England and Empress of India, dies 58 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes I. Kehat, D. Kenyagin-Karsenti, M. Snir, H. Segev, M. Amit, A. Gepstein, E. Livne, O. Binah, J. Itskovitz-Eldor, L. Gepstein J Clin Invest. 2001;108:407-414 E mbryonic stem (ES) cells from mice have revolutionized biomedical science since their isolation in 1981. Manipulation of murine ES cells has allowed the creation of transgenic and knockout mice that have greatly expanded our understanding of development and gene function on an organism level. The fundamental property that makes ES cells unique is totipotency, the ability to give rise to any cell lineage in the body. In 1998, Thompson et al first described the generation of human ES cell lines derived from a blastocyst. The isolated human ES cells were shown to have the capacity to differentiate into the ectodermal, endodermal, and mesodermal lineages. Kehat et al were the first to demonstrate that human ES cells in culture can be differentiated into myocytes that possess characteristics that define cardiomyocytes. The ability to reliably differentiate human ES cells into cardiomyocytes in the laboratory will be a powerful tool in understanding human cardiogenesis. Moreover, these findings raise the possibility of using human ES cells to achieve myocardial repair. Proof-of-concept studies in the mouse have suggested that embryonic cardiomyocytes can be useful for cardiac repair after injury. Many questions still need to be answered. The signals and events that promote human ES cell differentiation into cardiomyocytes are incompletely understood as only 8% of embryonic bodies exhibited contracting areas. By understanding these signals, human ES cells may be directed toward cardiomyocyte differentiation to increase the yield. The interaction between mature cardiomyocytes in a diseased heart and the ES cell–derived cardiomyocytes need to be understood as well as the processes that promote integration of the transplanted cardiomyocyte into the heart. Furthermore, issues related to rejection will have to be addressed with transplanted ES cell–derived cardiomyocytes before widespread clinical use. In this important manuscript, the authors allowed human ES cells to aggregate and form embryonic bodies containing derivatives of all three germ layers. The embryonic bodies were plated on gelatin-coated dishes and were observed daily to assess the presence of spontaneous contractions. Contracting areas were mechanically dissected from the embryonic bodies and rigorously studied. By using reverse transcriptase polymerase chain reaction (RT-PCR), the contracting area cells were found to express cardiac specific genes such as troponin I, troponin T, the transcription factors GATA4 and Nkx2.5, as well as atrial and ventricular myosin light chains. Contracting area cells exhibited strong immunostaining to cardiac myosin heavy chain, troponin I, ANP, α-actinin and desmin. Contractile elements ranging from unorganized myofibrillar bundles to organized sarcomeres and Z bands could be appreciated by electron microscopy. Extracellular electrograms of the contracting area cells demonstrated depolarization and repolarization activity. Positive and negative chronotropic responses were observed with the application of the β-adrenergic agonist isoproterenol and the muscarinic agonist carbamylcholine. The authors conclusively showed that the contracting areas cells possessed the gene expression profile, ultrastructure, immunoreactivity, and functional properties of human cardiomyocytes. 2001/1901 Jean Henri Dunant receives the Nobel Peace Prize for his role in founding the International Committee of the Red Cross; New York becomes the first state to make automobile license plates compulsory; and German psychiatrist Alois Alzheimer describes his eponymous disease 59 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Biological pacemaker created by gene transfer J. Miake, E. Marban, H. B. Nuss Nature. 2002;419:132-133 W hen gene therapy first entered medical scientific consciousness, clinical applications were focused on the cure of diseases resulting from defective or missing genes. By replacing deleterious genes with normal functional copies, disease can be halted or even reversed. Miake et al took a radical departure from this paradigm and showed that gene transfer can tweak existing cells to change their physiological role. The authors asked, “Can ventricular myocytes be engineered into pacemaker cells?” noted to “march through” the sinus beats and at times faster than sinus rhythm. Though the techniques are not clinically acceptable, this report clearly demonstrated for the first time that biological pacemakers are achievable. Furthermore, the findings shed insights into the electrophysiological makeup of ventricular myocytes. A particularly attractive aspect is that a biological pacemaker can be created from one’s own cardiac cells by ex vivo gene transfer. Ventricular myocytes can be harvested with a cardiac biotome, transduced with the gene of choice and reimplanted into the heart. Alternatively, the gene transfer vector can be injected directly into the myocardium. Miake et al’s findings triggered the race to develop a reliable biological pacemaker, with groups worldwide utilizing different viral vectors, cell-based delivery, and novel biomaterials. Obviously, many hurdles remain before clinical acceptance, especially regarding safety and reliability, and of course, proof that biological pacemakers are better than the current gold standard, the electronic pacemaker. Miake et al hypothesized that adult ventricular myocytes had the appropriate repertoire of ion channels for spontaneous pacemaker activity, but that this was normally repressed by the inward-rectifier potassium current (IK1) encoded by the Kir2 gene family. IK1 is not found in nodal pacemaker cells, but is robust in adult atrial and ventricular myocytes, where it stabilizes a very negative resting potential and suppresses excitability. Because Kir2 potassium channel genes have a tetrameric structure, a dominant negative strategy to suppress IK1 is feasible with a nonfunctional Kir2.1 mutant, in this case Kir2.1AAA, which has 3 alanine substitutions in the pore region. The dominant negative mutant was packaged with green fluorescent protein (GFP) into an adenoviral vector and introduced into the guinea pig left ventricle during transient cross-clamping of the great vessels. Gene transduction rates into ventricular myocytes were about 20% as seen by GFP expression and whole-cell recording of isolated GFP expressing myocytes had 80% suppression of IK1. The electrophysiology of the gene transduced myocytes fell into two categories: (i) no spontaneous activity, but prolonged elicited action potentials; or (ii) spontaneous activity remarkably similar to sinoatrial pacemaker cells. The myocytes with spontaneous activity had IK1 suppressed to a greater extent. The surface ECG of the transfected animals was fascinating. Half of the guinea pigs remained in sinus rhythm with QT prolongation. However, the other half had cardiac rhythms indicating spontaneous ventricular foci suggested by the broad QRS duration. The ventricular rhythms were 2002/1902 Mount Pelée (Montagne Pelée) volcano erupts in Martinique, spewing out a pyroclastic cloud that claimed more than 30 000 dead; St Marks’ Campanile collapses in Venice on 14th July—reconstruction was decided the same day by the Communal Council; and Indian mystic Swami Vivekananda dies, aged 39 60 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Heart regeneration in zebrafish K. D. Poss, L. G. Wilson, M. T. Keating Science. 2002;298:2188-2190 A apical amputation resulted in scar formation rather than cardiac regeneration, similar to the human cardiac injury response. fter a myocardial infarction, human hearts respond to the injury by extensive scarring with minimal regenerative potential. Replacement of myocardium with scar tissue has consequences for ventricular remodeling, cardiac function, and arrhythmia potential. Previous work has shown that the zebrafish (Danio rerio) is capable of regenerating fins, retina, and spinal cord. Poss and colleagues are the first to demonstrate that zebrafish can regenerate ventricular myocardium after acute injury. Stem cell–based approaches to cardiac repair have gotten a lot of attention, but an alternative strategy is to stimulate the damaged heart to heal itself. Though the zebrafish heart is simpler in structure than mammalian hearts, elucidating the signals and pathways that direct injured hearts toward regeneration or scar formation may give insights into human cardiac biology and strategies for treatment. Amphibians have also been shown to have the capacity for cardiac regeneration. However, cardiac regeneration would be easier to study in the zebrafish because of its sequenced genome and the ease of conducting mutational studies. Poss et al have identified a species that possesses robust cardiac regeneration capacity after injury with well-established genetics. The great promise of understanding cardiac regeneration in the fish is that it may lead to therapeutics that can stimulate cardiac regeneration in the injured human heart. Let us hope that we can fish out these factors. Zebrafish have become a favored species in genetic research due to their ease of handling, fast generation times, and availability of simple genetic screening methods. Using adult zebrafish, the heart was exposed through a small skin incision and the ventricular apex excised with scissors, about 20% of the heart. Profuse bleeding from the ventricular cavity was stopped with a piece of laboratory paper tissue and a large clot of erythrocytes formed over the excision site. A survival rate of 90% was achieved when 20% of the ventricle was excised. Mortality rates increased when more than 20% of the ventricle was removed. The zebrafish were then followed for up to 60 days after surgery. Within 2 to 4 days after ventricular apical amputation, fibrin began to replace the erythrocyte clot. The zebrafish during this time appeared sluggish, but by 1 week after amputation, they were indistinguishable from sham controls. Nine to 30 days after amputation, cardiac myofibers surrounded, penetrated, and eventually replaced the fibrin clot. By 60 days after amputation, the hearts that underwent ventricular apical amputation appeared grossly normal in size and shape. The zebrafish ventricle is composed of two myocardial layers, an outer compact layer, and an inner trabecular layer. The amputated hearts regenerated both myocardial layers and were indistinguishable on histological inspection. Using BrdU, a marker of DNA synthesis, Poss et al showed that the cardiomyocytes closest to the cut edge underwent cell division to replace the lost cardiomyocytes. A mitotic checkpoint kinase (mps1) is known to be required for zebrafish fin regeneration as well as cell proliferation. Using a conditional mps1 mutant zebrafish line, ventricular 2002/1902 Birth of Leni Riefenstahl, the German film director who shot the controversial “Triumph of the Will” propaganda film at the 1934 Nuremberg Congress of the Nazi Party; Edward VII is crowned King of the United Kingdom; and the Carnegie Institution is founded in Washington DC, in support of scientific research 61 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Adult cardiac stem cells are multipotent and support myocardial regeneration A. P. Beltrami, L. Barlucchi, D. Torella, M. Baker, F. Limana, S. Chimenti, H. Kasahara, M. Rota, E. Musso, K. Urbanek, et al Cell. 2003;114:763-776 E arlier work by these authors dispelled the long held belief that the adult human heart is incapable of cell division. The origins of the dividing cardiomyocytes were unclear. One intriguing possibility was a pool of resident cardiac stem cells that can be activated to proliferate and differentiate into cardiac cells. Beltrami et al demonstrate in this paper that adult mammalian hearts have such a defined pool of stem cells that can be isolated, cultured, and implanted into a myocardial infarct to regenerate myocardium and improve cardiac function. differentiated counterparts. The observed changes in vivo were not due to fusion of stem cells with native cardiac cells, as >99% of the cells examined were diploid, not tetraploid. More recently, these cardiac stem cells were delivered via the coronary circulation in rats with myocardial infarctions, with similar results. Beltrami et al are the first group to have defined a population of resident cardiac stem cells that were clonigenic, multipotent, and able to participate in the formation of functional myocardium within a clinical relevant model. If similar populations of cardiac stem cells are present in humans, the implications for clinical applications are obvious. An interesting question is that if a resident pool of cardiac stem cells is present, why do these cells not repair the myocardium efficiently after injury? One potential answer may be that a critical number of cardiac stem cells is needed for meaningful repair. The authors do not comment on dose response, if any, of their cardiac stem cells. Three other populations of cardiac stem cells have been described since Beltrami et al. Stay tuned for more developments in this rapidly evolving area. In adult rats, Beltrami et al characterized cardiac stem cells by the cell surface marker profile (c-Kit+, Lin-, CD45-, CD34-). These cardiac stem cells can be isolated from rat heart preparation by flow cytometry using FACS or magnetic beads coated with c-Kit antibody. The isolated cells can be expanded in culture indefinitely and cloned. In laboratory cultures, the cardiac stem cells were able to differentiate into cardiomyocytes, smooth muscle cells as well as endothelial cells, however in immature forms. For example, the culture differentiated cardiomyocytes that expressed specific markers such as α–actin and cardiac myosin heavy chain, but exhibited disorganized structures rather than sarcomeres, and spontaneous contraction was absent. To test the ability of these cardiac resident stem cells in myocardial repair, adult rats with induced myocardial infarction received injections of stem cells labeled with BrdU along the infarct border. After 10 days, a thin regenerating band was seen that incompletely penetrated the infarct. After 20 days, the entire infarct demonstrated BrdU labeled cells and a significant increase in myocardial volume. The infarct size was significantly decreased with stem cell treatment (70% control vs 48% stem cell) and ejection fraction was improved (34% control vs 45% stem cell). The labeled resident stem cells gave rise to cardiomyocytes, smooth muscle cells, and endothelial cell in the infarct border zone. In contrast to the culture differentiated cells, the labeled cells differentiated in the rat heart morphologically appeared mature. Isolated cardiomyocytes derived from stem cells had similar contractile function as native cardiomyocytes when tested in vitro, unlike their cultured 2003/1903 King Edward VII is proclaimed Emperor of India; the Martha Washington Hotel, exclusively reserved for women, is founded in New York; and American frontierswoman Calamity Jane dies, aged 51 62 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers I. Potapova, A. Plotnikov, Z. Lu, P. Danilo Jr, V. Valiunas, J. Qu, S. Doronin, J. Zuckerman, I. N. Shlapakova, J. Gao, et al Circ Res. 2004;94:952-959 T he electronic pacemaker is undoubtedly one of the major medical advances in history. Though highly successful, there is room for improvement, as electronic pacemakers have limited battery life, lack of autonomic response, and imply the presence of permanent hardware in the body. With regard to biological pacemakers, early efforts have focused on gene-based approaches utilizing different viral vectors. However, viral gene transfer raises questions concerning duration and magnitude of gene expression, the consequences of viral protein expression, carcinogenic, and infectious potential. With this paper, Potapova et al lay the foundation for using stem cell–based approaches to generate reliable cardiac pacemaking. site than controls. hMSC were easily identified histologically by their size and confirmed by positive vimentin and CD44 staining. Staining for Cx43 revealed that gap junctions formed between the hMSC and canine ventricular myocytes in vivo. No evidence of inflammation or rejection was seen, underscoring the immunoprivileged status of hMSC. More recent work by Plotinikov et al (Circulation 2007;116:706-713) has shown that HCN2-hMSC can provide reliable biological pacing for up to 6 weeks without rejection. Potapova et al were the first to show the feasibility of a stem-cell–based approach to biological pacemaker development. hMSC appear to have many desirable features of a delivery platform that may allow for widespread clinical use. Unlike viral-based approaches where reliable expression can be challenging from subject to subject, a stem cell–based pacemaker can be verified for expression and performance prior to implantation. The immunoprivileged status of hMSC may allow for an inventory of ready-to-use biological pacemakers without significant rejection. Many questions need to be answered before clinical testing of a biological pacemaker to compete against current electronic pacemakers. This important work brings us one beat closer. Human mesenchymal stem cells (hMSC) have several advantageous features making them attractive delivery vehicles. hMSC are readily available due to easy harvesting and can be maintained in culture. hMSC also possess local immunosuppressive properties that allow allogenic transplant without significant rejection, a feature that may ease clinical application. Potapova et al show that a robust If current is present in transfected hMSC with the mouse HCN2 gene by electroporation. Moreover, application of the β-adrenergic agonist isoproterenol induced a positive shift in If activation in the HCN2-hMSC. Acetylcholine, a muscarinic agonist, reversed the effects of isoproterenol. Therefore, HCN2-hMSC possesses the protein machinery required to respond to autonomic hormones. A pacemaker cell must electrically couple to neighboring cardiomyocytes to pace, and the investigators elegantly showed that hMSC do indeed form functional gap junctions. In cocultures of HCN2-hMSC with canine ventricular myocytes, dual whole-cell recording of hMSC and myocyte pairs demonstrate electrical coupling. Furthermore, ventricular myocytes cocultured with HCN2-hMSC have more positive maximum diastolic potentials and faster spontaneous rates than myocytes cultured with hMSC expressing GFP. The pacemaking performance of HCN2-hMSC was very impressive with implanted dogs having significantly faster idioventricular rates originating from the implant 2004/1904 Theodore Roosevelt is reelected President of the USA; Ivan Petrovich Pavlov is awarded the Nobel Prize in Physiology or Medicine; and Birth of Umm Kulthum (Oum Kalsoum), who was to be unanimously celebrated as a singer in the Arab world 63 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Regenerating the heart M. A. Laflamme, C. E. Murry Nat Biotechnol. 2005;23:845-856 W e have all been taught that the human heart is an end organ without any regenerative properties. Patients with failing hearts may receive a mechanical ventricular assist device or a heart transplant as treatment. Ventricular assist devices have a host of drawbacks such as infections, thromboembolic complications, and arrhythmias, limiting their chronic widespread use. Improved immunosuppressive regimens have made cardiac transplant a long-term solution for heart failure patients. However, there are simply not enough hearts available for the huge transplant demand. Advances in basic science over the past decade have initiated excited discussions on what used to be considered science fiction, “How to grow a new heart?” Laflamme and Murry have written an outstanding comprehensive review on the current state of progress in cardiac regeneration. development. At last count, four separate populations of resident myocardial progenitor cells have been described. Embryonic stem cells have the greatest hope of generating an entire heart, but significant challenges remain on how to coax them toward cardiogenesis. Tissue engineering will almost certainly play a major role in regenerative cardiac repair. The most common approaches utilize cell scaffolds, mechanical conditioned gels, and layered cellular sheets. One of the biggest challenges with tissue engineering is nutrient delivery, since diffusion alone can only supply to a depth 150 microns. Several obstacles also remain that limit widespread clinical applications. Cell delivery systems are less than ideal as the great majority of cells are lost in the circulation or leakage from injection site. Cell survival is a major problem, as most transplanted cells do not survive. Control of proliferation is difficult, as a delicate balance exists between cellular replacement and neoplasia. The underlying hypothesis is that heart failure can be reversed or prevented if new myocardium can be grown and integrated into diseased hearts. Early work toward cellbased cardiac repair utilized skeletal myoblasts injected into the heart; however, it became quickly clear that skeletal myoblasts remained skeletal, did not transdifferentiate into cardiomyocytes and did not electromechanically couple to the surrounding myocardium. Clinical transplantation studies provided some evidence that circulating cells had the ability to repopulate adult cardiac tissue, the most common case being a male patient receiving a female donor heart, with Y chromosome cells subsequently seen within the transplanted heart. Initial focus was on hematopoietic stem cells. The contribution of bone marrow stem cells to the healing myocardium was controversial, with several groups reporting conflicting results. Mesenchymal stem cells long thought to be permanent residents of the bone marrow stromal component have generated significant interest for therapeutic applications due to two fascinating properties. Mesenchymal stem cells appear to have local immunosuppressive properties that allow them to survive in allogenic settings and they appear to home to areas of injury. The discovery of resident myocardial progenitor cells in the adult heart changed what we understood about The field is moving very rapidly but it is highly unlikely that we will learn how to mend broken hearts soon. Nonetheless, phase 1 clinical testing has been initiated for several stem cell–based therapies for ischemic heart disease and, thus far, they appear to be safe and well tolerated. The medical community will have to wait for larger randomized multicenter trial results to gauge therapeutic value. 2005/1905 Tsar Nicholas II of Russia agrees to reintroduce an elected council, the Duma; Albert Einstein’s “miracle year,” which saw the four major publications that were to profoundly change the face of physics; and French novelist Jules Verne, author of “Twenty Thousand Leagues Under the Sea,” dies, aged 77 64 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction D. L. Kraitchman, M. Tatsumi, W. D. Gilson, T. Ishimori, D. Kedziorek, P. Walczak, W. P. Segars, H. H. Chen, D. Fritzges I. Izbudak, et al Circulation. 2005;112:1451-1461 T he human heart has limited regenerative capacity after a myocardial infarction (MI), a fact cardiologists are reminded of daily. Basic research has established the capability of stem cells to differentiate into cardiomyocytes. In animal models of myocardial injury, stem cell–based strategies have improved myocardial function. A crucial issue for any therapeutic is delivery to the target tissue. Histological examinations of postmortem cardiac tissue suggest that intravenously administered stem cells may preferentially localize to areas of injury. Kraitchman et al provide convincing data in a clinically relevant model of acute MI that mesenchymal stem cells (MSC) do indeed home, and furthermore stay, in the infarct region. to 7 days after injection showed diffuse myocardial uptake corresponding to the anterior apical infarct area, but not in normal myocardium. MRI imaging could not visualize the Feridex-labeled MSC due to the diffuse nature of the distribution. Postmortem histological examination confirmed the presence of the labeled MSC in the infarct and peri-infarct cardiac regions. Kraitchman et al have provided the methods for noninvasive tracking of MSC inside the living body. Translation into humans should be straightforward as the scanners and labels used are approved by the US Food and Drug Administration and are available at most medical centers. Localizing and quantifying the number of MSC to the infarct region will be essential in determining the appropriate clinical dose amount and schedule. Interpretation of clinical data will be enhanced as outcomes can be related to the number of MSC targeted to the infarct area. Though validation studies will be required using human MSC, this work is a major step toward properly conducted stem cell trials pertaining to cardiac repair. The study utilized two common cardiac imaging techniques, single photon emission computed tomography (SPECT)/CT and magnetic resonance imaging (MRI). Nontransmural MI was created in dogs by 90-minute balloon occlusion followed by reperfusion. Allogenic canine MSC dual-labeled with 111In oxine and Feridex was injected intravenously 3 days later. 111In oxine is used routinely to label leukocyte and its half-life (67.3 hours) allows for prolonged serial imaging by SPECT. In vitro assay with 111In oxine had no appreciable effect on MSC proliferation, viability or differentiation. SPECT/CT images were obtained on day of injection, 24 hours after injection, and up to 1 week afterwards. MRI images were obtained only with the final SPECT/CT scan. SPECT/CT permits high-resolution detection of the radiolabelled MSC with anatomic localization. Immediately after injection of labeled MSC, lung uptake predominated, with smaller amounts of uptake in the liver and kidney. Presumably, the relative large size of MSC (about 25 μm) may cause some difficulty traversing the pulmonary circulation. Twenty-four hours later, the radiodistribution was dramatically different as lung uptake is much lower and the predominant uptake is within the liver and spleen, suggesting redistribution to the reticuloendothelial system. The extracardiac distribution pattern was seen in both MI dogs and noninfarct controls. In the infarcted heart almost immediately after MSC injection, increased radiolabel was appreciated in the anterior apex. SPECT/CT imaging at 4 2005/1905 An earthquake in India kills more than 20 000; the US Army begins work on the Panama Canal; and the FIFA (International Federation of Association Football) is created, still going strong a century later 65 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Molecular ablation of ventricular tachycardia after myocardial infarction T. Sasano, A. D. McDonald, K. Kikuchi, J. K. Donahue Nat Med. 2006;11:1256-1258 V entricular tachycardia (VT) is unfortunately a common and often fatal complication of ischemic heart disease. Implantable cardiac defibrillators (ICD) have greatly improved survival. However, ICDs have their share of shortcomings, including inappropriate painful shocks, with their associated psychological consequences, as well as the fact that hardware is permanently present within the body. Current antiarrhythmic drugs are limited by incomplete arrhythmia suppression and toxicities, including proarrhythmic effects. Gene-based approaches to control arrhythmias are of great interest because they offer several distinct advantages. Expression vectors can be precisely delivered to the area of interest, such as the infarct border zone, to minimize systemic side effects with existing available technology (coronary catheterization or percutaneous endocardial injection). Unlike drugs that can only modulate ion channels and receptors expressed in the diseased cardiomyocyte, gene therapy is not limited by the existing protein repertoire, but can deliver any protein. The therapeutic protein can be an endogenous protein, a mutant, a chimera, or even a protein completely foreign to the cardiomyocyte. 12-lead ECG showed no difference among the three groups of pigs, including with respect to the QT interval. However, monophasic action potential duration and the effective refractory period were increased only in the anterior septum (gene transfer zone), but not in other areas of the heart of the G628S pigs. Patch clamping of isolated myocytes from the anterior septum of the G628S animals also exhibited prolonged action potential durations. Furthermore, gene transfer did not appear to be proarrhythmic, as spontaneous ventricular arrhythmias were not observed in any of the pigs over the 4 weeks of study. To compare against current antiarrhythmics, 3 pigs with infarcts were treated with dofetilide, a known KCNH2-blocking drug. Unlike the G628S pigs, dofetilide increased the QT interval, prolonged the ERP globally, and the pigs still had inducible VT. In this proof-of-concept study, Sasano et al were the first to demonstrate effective arrhythmia suppression with a gene transfer approach in a clinical relevant model. Sasano et al elegantly showed how local gene transfer via the coronary arteries is safe, effective, and can be a tailored therapeutic approach. Significant work lies ahead before clinical application is a reality. The adenovirus vector used in this study has short-lived expression of the order of 1 to 3 weeks. Long-term expression is required for clinical use. Stem cell–based gene delivery approaches are also promising. Sasano et al gave the first demonstration of a gene therapy approach to effectively suppress postinfarction VT. Myocardial infarctions were created in pigs by balloon occlusion of the mid-left anterior descending artery (LAD) for 150 minutes. After 3 weeks of recovery, VT inducibility was assessed by programmed stimulation and monomorphic VT was inducible in all pigs tested. Adenovirus expressing a dominant negative version of the KCNH2 (hERG) potassium channel (G628S) was then locally infused into the midLAD with a catheter (the same site as for infarction balloon occlusion). KCNH2 potassium currents are involved in repolarization. Numerous drugs that block KCNH2 prolong the QT interval and are proarrhythmic. In all pigs treated with G628S, VT was no longer inducible. Two other groups receiving saline or adenovirus expressing the lacZ-reporter gene continued to have inducible VT. Sinus intracardiac electrograms in all pigs showed low amplitude fractionated electrical activation within the gene transfer zone. Surface 2006/1906 The car manufacturing company Rolls-Royce Ltd is founded by Henry Royce and Charles Stewart Rolls; the first Victrola record player, a ponderous machine enclosed in a wooden cabinet, is manufactured in the USA; and SOS (later associated with the phrase Save Our Souls) becomes the first internationally recognized distress signal 66 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Summaries of Ten Seminal Papers - Lau Theoretical impact of the injection of material into the myocardium: a finite element model simulation S. T. Wall, J. C. Walker, K. E. Healy, M. B. Ratcliffe, J. M. Guccione Circulation. 2006;114:2627-2635 S tem cell transplantation by direct injection into the myocardial infarction area has gained significant attention as a strategy to improve cardiac function and prevent clinical heart failure. Numerous preclinical experiments and several small clinical trials using stem cell–based therapies have shown small, but significant, improvement in cardiac function after treatment. However, convincing evidence that stem cell–derived cardiomyocytes working in concert with the native myocardium as the underlying reason for functional improvement has been absent. Cellular elements derived from implanted stem cells are found within the infarct area, but their numbers are quite small in comparison with the magnitude of cardiac functional improvement. Wall and colleagues were the first to question whether the improvement in cardiac function after stem cell injection into the infarct heart was due to the passive mechanical consequences of the injection rather than to the stem cells. Injections in the infarct zone improved ejection fraction as well as the stroke volume/end-diastolic volume (SV/EDV) relationship, but did not affect the SV/EDP (end-diastolic pressure) relationship. An obvious study limitation is the reliance of the results on the accuracy of the computational model in reflecting an infarcted left ventricle. Furthermore, the simulations only model the immediate consequence of an injection volume, the long-term consequences are not predicted by this model. Nonetheless, Wall et al were among the first to rigorously examine the passive properties of cardiac injections on function. An intracardiac injection of a cellular therapeutic must be suspended in a solution or a biomaterial. The biomedical community has placed enormous emphasis on the cellular elements, but largely ignored the medium carrying the cells. Cell therapy efficacy must separately evaluate active cellular contributions and passive mechanical contributions. It is becoming quite clear that the future of cardiac repair will require a harmonious marriage of cellular technology with advanced biomaterials. The experimental protocols used to inject stem cells into the heart may have significant mechanical consequences. The authors point out that recent rat experiments used 50 μL of fibrin gel with stem cells to inject into the left ventricle. The average heart mass of an adult rat is about 1 g, so a 50-μL injection amounts to 5% of total heart mass. Experiments in mice are even more exaggerated, as a 50-μL injection into the heart would correspond to 50% of heart mass (average adult mice heart mass is 100 mg). In humans, in the Bone Marrow Transfer to Enhance ST-Elevation Infarct Regeneration (BOOST) trial, a 26-mL injection of stem cells was introduced into the infarct area. With the average human left ventricular wall volume being about 300 mL, this is over 8% of LV wall volume. Wall et al used a 216-element mesh computational model of a sheep left ventricle with an anterior apical infarct to calculate the effect of materials of varying stiffness and volume on wall stress and cardiac function. They looked at the effects of a single infarct border zone injection, multiple border zone injections, and injections into the infarct. They conclude that injections in the border zone decrease endsystolic fiber stress in proportion to the volume injected. 2006/1906 Mount Vesuvius erupts, devastating the city of Naples; Mahatma Gandhi adopts nonviolence as a form of political resistance in South Africa; and Norwegian playwright Henrik Ibsen dies, aged 78 67 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Mending the Broken Heart Bibliography of One Hundred Key Papers selected by Ira S. Cohen*†, MD, PhD and Glenn R. Gaudette ‡, PhD * Department of Physiology and Biophysics - †Institute for Molecular Cardiology - Stony Brook University - NY ‡ Department of Biomedical Engineering - Worcester Polytechnic Institute - Worcester - Mass - USA Tumor stromal-derived factor-1 recruits vascular progenitors to mitotic neovasculature, where microenvironment influences their differentiated phenotypes. Aghi M, Cohen KS, Klein RJ, Scadden DT, Chiocca EA. Cancer Res. 2006;66:9054-9064. Cell therapy in myocardial infarction. Arnesen H, Lunde K, Aakhus S, Forfang K. Lancet. 2007;369:2142-2143. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. Nature. 2004;428:668-673. Inhibitory G protein overexpression provides physiologically relevant heart rate control in persistent atrial fibrillation. Bauer A, McDonald AD, Nasir K, et al. Circulation. 2004;110:3115-3120. Adult cardiac stem cells are multipotent and support myocardial regeneration. Beltrami AP, Barlucchi L, Torella D, et al. Cell. 2003;114:763-776. Evidence that human cardiac myocytes divide after myocardial infarction. Beltrami AP, Urbanek K, Kajstura J, et al. N Engl J Med. 2001;344:1750-1757. Mesenchymal stem cell injection after myocardial infarction improves myocardial compliance. Berry MF, Engler AJ, Woo YJ, et al. Am J Physiol Heart Circ Physiol. 2006;290:H2196-H2203. Cardiac HCN channels: structure, function, and modulation. Biel M, Schneider A, Wahl C. Trends Cardiovasc Med. 2002;12:206-212. Cyclin D2 induces proliferation of cardiac myocytes and represses hypertrophy. Busk PK, Hinrichsen R, Bartkova J, et al. Exp Cell Res. 2005;304:149-161. Stem cells: lost in translation. Chien KR. Nature. 2004;428:607-608. Creation of a biological pacemaker by cell fusion. Cho HC, Kashiwakura Y, Marban E. Circ Res. 2007;100:1112-1115. Allogeneic mesenchymal stem cell transplantation in postinfarcted rat myocardium: short- and long-term effects. Dai W, Hale SL, Martin BJ, et al. Circulation. 2005;112:214-223. Copyright © 2009 LLS SAS. All rights reserved 69 www.dialogues-cvm.org Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Bone marrow-derived cardiomyocytes are present in adult human heart: a study of gender-mismatched bone marrow transplantation patients. Deb A, Wang S, Skelding KA, Miller D, Simper D, Caplice NM. Circulation. 2003;107:1247-1249. The contribution of the “pacemaker” current (I f) to generation of spontaneous activity in rabbit sino-atrial node myocytes. DiFrancesco D. J Physiol. 1991;434:23-40. Direct activation of cardiac pacemaker channels by intracellular cyclic AMP. DiFrancesco D, Tortora P. Nature. 1991;351:145-147. Cell-based therapy of myocardial infarction. Dimmeler S, Burchfield J, Zeiher AM. Arterioscler Thromb Vasc Biol. 2008;28:208-216. Aging and disease as modifiers of efficacy of cell therapy. Dimmeler S, Leri A. Circ Res. 2008;102:1319-1330. Focal modification of electrical conduction in the heart by viral gene transfer. Donahue JK, Heldman AW, Fraser H, et al. Nat Med. 2000;6:1395-1398. Gene therapy for cardiac arrhythmias. Donahue JK, Kikuchi K, Sasano T. Trends Cardiovasc Med. 2005;15:219-224. Cell cycle regulation to repair the infarcted myocardium. Dowell JD, Field LJ, Pasumarthi KB. Heart Fail Rev. 2003;8:293-303. Enhancement of murine cardiac chronotropy by the molecular transfer of the human beta 2 adrenergic receptor cDNA. Edelberg JM, Aird WC, Rosenberg RD. J Clin Invest. 1998;101:337-343. p38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes. Engel FB, Schebesta M, Duong MT, et al. Genes Dev. 2005;19:1175-1187. Cardioprotective c-kit+ cells are from the bone marrow and regulate the myocardial balance of angiogenic cytokines. Fazel S, Cimini M, Chen L, et al. J Clin Invest. 2006;116:1865-1877. Multiply attenuated, self-inactivating lentiviral vectors efficiently deliver and express genes for extended periods of time in adult rat cardiomyocytes in vivo. Fleury S, Simeoni E, Zuppinger C, et al. Circulation. 2003;107:2375-2382. Rescue of cardiac defects in id knockout embryos by injection of embryonic stem cells. Fraidenraich D, Stillwell E, Romero E, et al. Science. 2004;306:247-252. Direct in vivo gene transfer into porcine myocardium using replication-deficient adenoviral vectors. French BA, Mazur W, Geske RS, Bolli R. Circulation. 1994;90:2414-2424. Gene therapy for myocardial infarction–associated congestive heart failure: how far have we got? Hammond HK, Tang T. Dialogues Cardiovasc Med. 2009;14:27-34. 70 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Human embryonic stem cells develop into multiple types of cardiac myocytes: action potential characterization. He JQ, Ma Y, Lee Y, Thomson JA, Kamp TJ. Circ Res. 2003;93:32-39. Radiolabeled cell distribution after intramyocardial, intracoronary, and interstitial retrograde coronary venous delivery: implications for current clinical trials. Hou D, Youssef EA, Brinton TJ, et al. Circulation. 2005;112:I150-I156. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Hsieh PC, Segers VF, Davis ME, et al. Nat Med. 2007;13:970-974. Suppression of electrical alternans by overexpression of HERG in canine ventricular myocytes. Hua F, Johns DC, Gilmore RF Jr. Am J Physiol Heart Circ Physiol. 2004;286:H2342-H2352. Rate dependence and regulation of action potential and calcium transient in a canine cardiac ventricular cell model. Hund TJ, Rudy Y. Circulation. 2004;110:3168-3174. Gene transfer of a synthetic pacemaker channel into the heart: a novel strategy for biological pacing. Kashiwakura Y, Cho HC, Barth AS, Azene E, Marban E. Circulation. 2006;114:1682-1686. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. Kehat I, Kenyagin-Karsenti D, Snir M, et al. J Clin Invest. 2001;108:407-414. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Kehat I, Khimovich L, Caspi O, et al. Nat Biotechnol. 2004;22:1282-1289. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Kinnaird T, Stabile E, Burnett MS, et al. Circ Res. 2004;94:678-685. Tissue-engineered myocardial patch derived from extracellular matrix provides regional mechanical function. Kochupura PV, Azeloglu EU, et al. Circulation. 2005;112:I144-I149. Novel injectable bioartificial tissue facilitates targeted, less invasive, large-scale tissue restoration on the beating heart after myocardial injury. Kofidis T, Lebl DR, Martinez EC, Hoyt G, Tanaka M, Robbins RC. Circulation. 2005;112:I173-I177. Engraftment of engineered ES cell-derived cardiomyocytes but not BM cells restores contractile function to the infarcted myocardium. Kolossov E, Bostani T, Roell W, et al. J Exp Med. 2006;203:2315-2327. Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction. Kraitchman DL, Tatsumi M, Gilson WD, et al. Circulation. 2005;112:1451-1461. 71 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Laflamme MA, Chen KY, Naumova AV, et al. Nat Biotechnol. 2007;25:1015-1024. Regenerating the heart. Laflamme MA, Murry CE. Nat Biotechnol. 2005;23:845-856. Intracoronary adenovirus encoding adenylyl cyclase VI increases left ventricular function in heart failure. Lai NC, Roth DM, Gao MH, et al. Circulation. 2004;110:330-336. Epicardial border zone overexpression of skeletal muscle sodium channel, SkM1, normalizes activation, preserves conduction, and suppresses ventricular arrhythmia: an in silico, in vivo, in vitro study. Lau DH, Clausen C, Sosunov EA, et al. Circulation. 2009;119:19-27. Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Laugwitz KL, Moretti A, Lam J, et al. Nature. 2005;433:647-653. Heart regeneration in adult MRL mice. Leferovich JM, Bedelbaeva K, Samulewicz S, et al. Proc Natl Acad Sci U S A. 2001;98:9830-9835. Human embryonic stem cell transplantation to repair the infarcted myocardium. Leor J, Gerecht S, Cohen S, et al. Heart. 2007;93:1278-1284. Low-molecular-weight peptides derived from extracellular matrix as chemoattractants for primary endothelial cells. Li F, Li W, Johnson S, Ingram D, Yoder M, Badylak S. Endothelium. 2004;11:199-206. Overexpression of HCN-encoded pacemaker current silences bioartificial pacemakers. Lieu DK, Chan YC, Lau CP, Tse HF, Siu CW, Li RA. Heart Rhythm. 2008;5:1310-1317. A family of hyperpolarization-activated mammalian cation channels. Ludwig A, Zong X, Jeglitsch M, Hofmann F, Biel M. Nature. 1998;393:587-591. Growth factors mobilize CXCR4 low/negative primitive hematopoietic stem/progenitor cells from the bone marrow of nonhuman primates. Mahmud N, Patel H, Hoffman R. Biol Blood Marrow Transplant. 2004;10:681-690. Mesenchymal stem cells modified with Akt prevent remodeling and restore performance of infarcted hearts. Mangi AA, Noiseux N, Kong D, et al. Nat Med. 2003;9:1195-201. Improved left ventricular aneurysm repair with bioengineered vascular smooth muscle grafts. Matsubayashi K, Fedak PW, Mickle DA, Weisel RD, Ozawa T, Li RK. Circulation. 2003;108(suppl 1):II219-II225. Generation of functional murine cardiac myocytes from induced pluripotent stem cells. Mauritz C, Schwanke K, Reppel M, et al. Circulation. 2008;118:507-517. Current status and future prospects for cell transplantation to prevent congestive heart failure. Menasche P. Semin Thorac Cardiovasc Surg. 2008;20:131-137. 72 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers The Myoblast Autologous Grafting in Ischemic Cardiomyopathy (MAGIC) trial: first randomized placebo-controlled study of myoblast transplantation. Menasche P, Alfieri O, Janssens S, et al. Circulation. 2008;117:1189-1200. Autologous skeletal myoblast transplantation for severe postinfarction left ventricular dysfunction. Menasche P, Hagege AA, Vilquin JT, et al. J Am Coll Cardiol. 2003;41:1078-1083. Isolation and expansion of adult cardiac stem cells from human and murine heart. Messina E, De Angelis L, Frati G, et al. Circ Res. 2004;95:911-921. Biological pacemaker created by gene transfer. Miake J, Marban E, Nuss HB. Nature. 2002;419:132-133. Functional role of inward rectifier current in heart probed by Kir2.1 overexpression and dominant-negative suppression. Miake J, Marban E, Nuss HB. J Clin Invest. 2003;111:1529-1536. Stem cell therapy enhances electrical viability in myocardial infarction. Mills WR, Mal N, Kiedrowski MJ, et al. J Mol Cell Cardiol. 2007;42:304-314. Elastin stabilizes an infarct and preserves ventricular function. Mizuno T, Yau TM, Weisel RD, Kiani CG, Li RK. Circulation. 2005;112:I81-I88. Creation of a genetic calcium channel blocker by targeted gem gene transfer in the heart. Murata M, Cingolani E, McDonald AD, Donahue JK, Marban E. Circ Res. 2004;95:398-405. Regeneration gaps: observations on stem cells and cardiac repair. Murry CE, Reinecke H, Pabon LM. J Am Coll Cardiol. 2006;47:1777-1785. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Murry CE, Soonpaa MH, Reinecke H, et al. Nature. 2004;428:664-668. Directed and systematic differentiation of cardiovascular cells from mouse induced pluripotent stem cells. Narazaki G, Uosaki H, Teranishi M, et al. Circulation. 2008;118:498-506. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. Nussbaum J, Minami E, Laflamme MA, et al. FASEB J. 2007;21:1345-1357. Perfusion-decellularized matrix: using nature’s platform to engineer a bioartificial heart. Ott HC, Matthiesen TS, Goh SK, Black LD, Kren SM, Netoff TI, Taylor DA. Nat Med. 2008;14:213-221. Stem-cell-based therapy and lessons from the heart. Passier R, van Laake LW, Mummery CL. Nature. 2008;453:322-329. CD31- but Not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Pfister O, Mouquet F, Jain M, et al. Circ Res. 2005;97:52-61. 73 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Multilineage potential of adult human mesenchymal stem cells. Pittenger MF, Mackay AM, Beck SC, et al. Science. 1999;284:143-147. Mesenchymal stem cells and their potential as cardiac therapeutics. Pittenger MF, Martin BJ. Circ Res. 2004;95:9-20. Spontaneous cardiomyocyte differentiation from adipose tissue stroma cells. Planat-Benard V, Menard C, Andre M, et al. Circ Res. 2004;94:223-229. HCN212-channel biological pacemakers manifesting ventricular tachyarrhythmias are responsive to treatment with If blockade. Plotnikov AN, Bucchi A, Shlapakova I, et al. Heart Rhythm. 2008;5:282-288. Xenografted adult human mesenchymal stem cells provide a platform for sustained biological pacemaker function in canine heart. Plotnikov AN, Shlapakova I, Szabolcs MJ, et al. Circulation. 2007;116:706-713. Biological pacemaker implanted in canine left bundle branch provides ventricular escape rhythms that have physiologically acceptable rates. Plotnikov AN, Sosunov EA, Qu J, et al. Circulation. 2004;109:506-512. Heart regeneration in zebrafish. Poss KD, Wilson LG, Keating MT. Science. 2002;298:2188-2190. Enhanced recovery of mechanical function in the canine heart by seeding an extracellular matrix patch with mesenchymal stem cells committed to a cardiac lineage. Potapova I, Doronin S, Kelly D, et al. Am J Physiol Heart Circ Physiol. 2008;295:H2257-H2263. Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers. Potapova I, Plotnikov A, Lu Z, et al. Circ Res. 2004;94:952-959. Expression and function of a biological pacemaker in canine heart. Qu J, Plotnikov AN, Danilo P Jr, et al. Circulation. 2003;107:1106-1109. Ischemic tissue repair by autologous bone marrow-derived stem cells: scientific basis and preclinical data. Quraishi A, Losordo DW. Handb Exp Pharmacol. 2007:167-179. Mathematical analysis of canine atrial action potentials: rate, regional factors, and electrical remodeling. Ramirez RJ, Nattel S, Courtemanche M. Am J Physiol Heart Circ Physiol. 2000;279:H1767-H1785. Therapeutic opportunities for cell cycle re-entry and cardiac regeneration. Regula KM, Rzeszutek MJ, Baetz D, Seneviratne C, Kirshenbaum LA. Cardiovasc Res. 2004;64:395-401. CXCR4 and CCR5 mediate homing of primitive bone marrowderived hematopoietic cells to the postnatal thymus. Robertson P, Means TK, Luster AD, Scadden DT. Exp Hematol. 2006;34:308-319. Finding fluorescent needles in the cardiac haystack: tracking human mesenchymal stem cells labeled with quantum dots for quantitative in vivo three-dimensional fluorescence analysis. Rosen AB, Kelly DJ, Schuldt AJ, et al. Stem Cells. 2007;25:2128-2138. 74 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Genes, stem cells and biological pacemakers. Rosen MR, Brink PR, Cohen IS, Robinson RB. Cardiovasc Res. 2004;64:12-23. Gene and cell therapy for life-threatening cardiac arrhythmias: will they replace drugs, surgery, and devices? Rosen MR, Danilo P Jr, Robinson RB. Dialogues Cardiovasc Med. 2009;14:44-51. Identification of a gene encoding a hyperpolarization-activated pacemaker channel of brain. Santoro B, Liu DT, Yao H, et al. Cell. 1998;93:717-729. Molecular ablation of ventricular tachycardia after myocardial infarction. Sasano T, McDonald AD, Kikuchi K, Donahue JK. Nature Med. 2006;12:1256-1258. Pilot trial on determinants of progenitor cell recruitment to the infarcted human myocardium. Schachinger V, Aicher A, Dobert N, et al. Circulation. 2008;118:1425-1432. Improved clinical outcome after intracoronary administration of bone-marrow-derived progenitor cells in acute myocardial infarction: final 1-year results of the REPAIR-AMI trial. Schachinger V, Erbs S, Elsasser A, et al. Eur Heart J. 2006;27:2775-2783. Monocyte chemotactic protein-3 is a myocardial mesenchymal stem cell homing factor. Schenk S, Mal N, Finan A, et al. Stem Cells. 2007;25:245-251. Repairing damaged myocardium: evaluating cells used for cardiac regeneration. Schuldt AJ, Rosen MR, Gaudette GR, Cohen IS. Curr Treat Options Cardiovasc Med. 2008;10:59-72. Myocardial neovascularization by bone marrow angioblasts results in cardiomyocyte regeneration. Schuster MD, Kocher AA, Seki T, et al. Am J Physiol Heart Circ Physiol. 2004;287:H525-H532. Cell isolation procedures matter: a comparison of different isolation protocols of bone marrow mononuclear cells used for cell therapy in patients with acute myocardial infarction. Seeger FH, Tonn T, Krzossok N, Zeiher AM, Dimmeler S. Eur Heart J. 2007;28:766-772. Cytokine preconditioning promotes codifferentiation of human fetal liver CD133+ stem cells into angiomyogenic tissue. Shmelkov SV, Meeus S, Moussazadeh N, et al. Circulation. 2005;111:1175-1183. A tissue engineering approach to progenitor cell delivery results in significant cell engraftment and improved myocardial remodeling. Simpson D, Liu H, Fan TH, Nerem R, Dudley SC Jr. Stem Cells. 2007;25:2350-2357. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Smith RR, Barile L, Cho HC, et al. Circulation. 2007;115:896-908. Adeno-associated viral vector delivers cardiac-specific and hypoxiainducible VEGF expression in ischemic mouse hearts. Su H, Joho S, Huang Y, et al. Proc Natl Acad Sci U S A. 2004;101:16280-16285. 75 Dialogues in Cardiovascular Medicine - Vol 14 . No. 1 . 2009 Bibliography of One Hundred Key Papers Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Takahashi K, Tanabe K, Ohnuki M, et al. Cell. 2007;131:861-172. Bioartificial sinus node constructed via in vivo gene transfer of an engineered pacemaker HCN channel reduces the dependence on electronic pacemaker in a sick-sinus syndrome model. Tse HF, Xue T, Lau CP, et al. Circulation. 2006;114:1000-1011. Human mesenchymal stem cells make cardiac connexins and form functional gap junctions. Valiunas V, Doronin S, Valiuniene L, et al. J Physiol. 2004;555:617-626. Fibroblast growth factor-2 regulates myocardial infarct repair: effects on cell proliferation, scar contraction, and ventricular function. Virag JA, Rolle ML, Reece J, Hardouin S, Feigl EO, Murry CE. Am J Pathol. 2007;171:1431-1440. Theoretical impact of the injection of material into the myocardium: a finite element model simulation. Wall ST, Walker JC, Healy KE, Ratcliffe MB, Guccione JM. Circulation. 2006;114:2627-2635. Does cell therapy for myocardial infarction and heart failure work? Wollert KC, Drexler H. Dialogues Cardiovasc Med. 2009;14:35-41. Intracoronary autologous bone-marrow cell transfer after myocardial infarction: the BOOST randomised controlled clinical trial. Wollert KC, Meyer GP, Lotz J, et al. Lancet. 2004;364:141-148. Platelet-derived growth factor-AB promotes the generation of adult bone marrow-derived cardiac myocytes. Xaymardan M, Tang L, Zagreda L, et al. Circ Res. 2004;94:E39-E45. Collagen-GAG scaffolds grafted onto myocardial infarcts in a rat model: a delivery vehicle for mesenchymal stem cells. Xiang Z, Liao R, Kelly MS, Spector M. Tissue Eng. 2006;12:2467-2478. Functional integration of electrically active cardiac derivatives from genetically engineered human embryonic stem cells with quiescent recipient ventricular cardiomyocytes: insights into the development of cell-based pacemakers. Xue T, Cho HC, Akar FG, et al. Circulation. 2005;111:11-20. Clonally expanded novel multipotent stem cells from human bone marrow regenerate myocardium after myocardial infarction. Yoon YS, Wecker A, Heyd L, et al. J Clin Invest. 2005;115:326-338. Engineered heart tissue for regeneration of diseased hearts. Zimmermann WH, Melnychenko I, Eschenhagen T. Biomaterials. 2004;25:1639-1647. 76


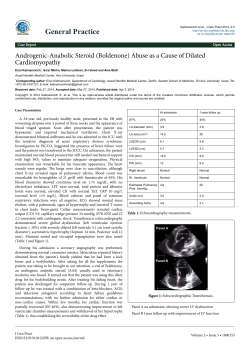








© Copyright 2026